| MR1 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Identifiers | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Aliases | MR1, HLALS, Major histocompatibility complex, class I-related | ||||||||||||||||||||||||||||||||||||||||||||||||||
| External IDs | OMIM: 600764 MGI: 1195463 HomoloGene: 123981 GeneCards: MR1 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Wikidata | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||





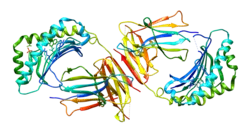
Major histocompatibility complex class I-related gene protein (MR1) is a non-classical MHC class I protein, that binds vitamine metabolites (intermediates of riboflavin synthesis) produced in certain types of bacteria. MR1 interacts with mucosal associated invariant T cells (MAIT).[5][6]
Gene location
MR1 is a protein that in humans is encoded by the MR1 gene and located on chromosome 1. Non-classical MHC class I genes are very often located on the same chromosome (mice chromosome 6, human chromosome 17) and interspaced within the same loci as the classical MHC genes. MR1 is located on another chromosome, the detailed gene analysis revealed that MR1 is a paralog originated by duplication of MHC locus on chromosome 6 (mice). This functional gene has been found in almost all mammals, proving the importance of MR1 across the mammalian kingdom and the fact that the duplication occurred early in the evolution of vertebrates.[5][7][8]
Another non-classical MHC class I CD1 is missing in certain species. There is 90% protein homology of the MR1 binding site between mice and humans. MR1 shares greater homology with classical MHC I class than with non-classical MHC class I. The human MR1 protein has 341 amino acid residues with a molecular weight of 39 366 daltons.[9][5]
Structure
MR1, like other MHC class I molecules, is composed of α1, α2 and α3 domains. α1 and α2 interact and together bind the antigen. The ligand binding pocket is small and contains aromatic and basic residues. Its small size limits it to only binding molecules of a similarly small size. α3 interacts with β2 microglobulin.[5]
Many different isoforms of MR1 have been identified. Many of the identified proteins have a premature terminating codon which generates non-functional proteins. MR1B isoform lacks the α3 domain. The α3 domain interacts with β2 microglobulin. This interaction and binding of the antigen stabilize the MHC I molecule. In the case of MR1B β2 microglobulin is not needed for stabilization of the structure. MR1B is expressed on the cell surface. This isoform binds antigen via α1 and α2 interaction. Some bacteria are able to target specific β2 microglobulin that enable MHC I presentation. This might be a mechanism used to avoid bacterial immune evasion during bacterial infections.[5]
Antigen presentation
The MR1 protein is capable of binding to molecules derived from bacterial riboflavin biosynthesis, and then present them to MAIT for activation.[7][8]
MR1 is almost undetectable under physiological conditions, surface expression increase in cells infected by microbes. Due to the antigen necessity for MR1 stabilization. MR1 binds the intermediates of riboflavine synthesis.
Many human cells can present antigens via MR1 with varying efficiency.[10][11][12] Human body can't synthesize most of the vitamins, thus the presence of intermediates of riboflavin synthesis is a marker of non-self. Many bacteria are capable of vitamine synthesis.[13] The first discovered MR1 ligand was 6-formyl pterin (6-FP).[5]
Within cells, MR1 is mostly stored inside the endoplasmic reticulum (ER), where ligand binding occurs.[14] Inside the ER, MR1 is stabilised in a ligand-receptive conformation by chaperone proteins tapasin and TAP-binding protein related (TAPBPR) to facilitate ligand binding.[14] After antigen binding MR1 undergoes conformational change, associate with β2 microglobulin and is directed to the cell membrane.[15][5][13]
MR1 stimulation is needed for MAIT development in thymus, the mechanism of antigen presentation in thymus is not clear.[5]
MR1 ligands
The MR1 pocket is composed primary of aromatic and basic amino acids and the volume is small, thus suitable for binding small molecule ligands.
The majority of MR1 ligands are uracil analogues, but some non-uracil drug-like molecules[16] also weakly bind to MR1.[17][18] The ligands 5-OP-RU and 5-OE-RU are compounds derived from riboflavin biosynthesis that bind MR1 for presentation to MAIT cells for activation.[7] They are chemically unstable, but have been synthesised as chemical tools for studying MR1 biology.[19] Ac-6-FP (acetyl-6-formylpterin) and 6-FP (6-formylpterin) also bind MR1, but they do not activate MAIT cells.[8] There is an evidence, that MR1 can bind other antigens. MR1 was able to stimulate T lymphocytes in the presence of Streptoccocus pyogenes, that is unable to synthesise riboflavin. MR1 is important in the immune fight against cancer, because MR1 T lymphocytes were able to selectively kill various cancer cells.[20]
Clinical significance
Cancer
Since the MR1 molecule is involved in presentation of cancer specific antigens and plays a role in tumor immunosurveillance, it has potential use in immunotherapy.[21]
Specific clones of MR1 T lymphocytes (MC.7.G5) were able to kill various cancer cells in vivo and in vitro and were inert to noncancerous cells. The MR1 expression on cancer cells is basal and appeared to be independent of bacterial load and MR1-ligand binding. Interestingly, cancer cell lines lacking a surface expression of MR1 weren't killed by MR1 T lymphocytes. Same result was shown in reaction to healthy but stressed or damaged cells, which were unable to activate MR1 T lymphocytes. This again suggests that some MR1 T lymphocytes can specifically react to a cancer cell derived ligand presented on the MR1 molecule.[22]
MAIT cells reactive to bacterial antigens are known as indirect tumor growth promoters with low cytotoxic activity. On the contrary, self-reactive MR1-restricted T cells (described above) not only promote inflammation and have much higher cytotoxic activity but also directly recognize tumor antigens presented on MR1 of cancer cells. This direct contact results in secretion of apoptosis-inducing factors and in death of a cancer cell.[23]
Taken together, immune response triggered by MR1-TCR interaction depends on the antigen presented on the MR1 receptor. As MAIT cells are enriched in mucosal sites like lungs or intestine, we can more likely expect a bacterial antigen presentation, which results in a different reaction of MAIT cells. It is shown that these cells inhibit NK cell and CD8+ T cell effector activity and the production of IFNγ in the response to bacterial antigens presented on MR1.[24] As a result, we should be careful when manipulating with MR1 molecule as a therapeutic target. On one hand its depletion can prevent unpreferable polarization of immune cells and impaired NK and CD8+ T cells activity. On the other hand, as described earlier, cancer cells lacking MR1 had better survival rate as they were not recognized by MR1 T lymphocytes (lower immune surveillance of cancer). To make MR1 molecule truly clinically significant we need to get a better understanding of mechanisms and differences in antigen presentation. In several studies it has been shown that MR1 uses more than one pathway to capture and traffic metabolite antigen depending on its source.[21]
Diabetes
As gut microbiota is modified in patients with diabetes,[25] it is expected that MR1-TCR interaction will have an impact on the disease progression.
MR1, as has been shown in some studies, plays an important role in promoting inflammation during obesity and Type 2 diabetes (T2D), specifically in adipose tissue and guts.[26] The study showed that mice lacking MR1 had a significantly decreased transcript level of cytokines/chemokines known to be associated with inflammation, such as Ccl2, Ccl5, Il1β, Il6, Il17a, Ifnγ, and Tnfα. In contrast, the transcript level of regulatory factors (Foxp3, Il5, and Il13) was increased.[26]
Situation is a bit different when it comes to Type 1 diabetes (T1D). Both in patients with T1D and in non-obese diabetic (NOD) mice an alteration in frequency and functions of MAIT cells was detected. On the other hand, NOD mice lacking MR1 had greater anti-islet autoreactive T cell response and local activation of dendritic cells which led to pancreatic islet destruction. These mice had overall exacerbated diabetes in comparison to the control group.[27]
Inflammatory bowel diseases (IBD)
Some studies show that MAIT cells infiltrate the colon in patients with ulcerative colitis. Haga et al. have previously reported that these cells also play an important role in pathogenesis of inflammatory bowel diseases.[28] An important role of MR1-TCR interaction in ulcerative colitis has been shown on MR1-deficient mouse model. Interestingly, both deficient and control mice developed colitis, though survival rate in deficient mice was much higher, than in control group. This deficiency decreased overall inflammation in the colon and reduced colitis severity.[29][30]
Arthritis
Both ankylosing spondylitis (AS) and rheumatoid arthritis (RA) are chronic inflammatory diseases affecting mostly bones and joints. Both of these diseases are also known as autoimmune or autoinflammatory diseases due to the presence of specific autoantibodies. It is also worth mentioning that they are often associated with other diseases such as IBD or psoriasis.[31] As microbiome is changed in both IBD and psoriasis, it is expected that MAIT cells will again play an important role in overall pathogenesis of all these conditions. In both AS and RA systemic frequency of MAIT cells was decreased.[32] On the contrary, it was highly elevated in synovial fluid. Also some phenotypical changes have been shown. For example, activation of MAIT cells positively correlated with AS progression in patients. There was also higher production of IL-17 by MAIT cells in peripheral blood and even higher in synovial fluid (none of these appeared in RA). Interestingly, it was observed only in male patients.[30][33][34]
References
- 1 2 3 GRCh38: Ensembl release 89: ENSG00000153029 - Ensembl, May 2017
- 1 2 3 GRCm38: Ensembl release 89: ENSMUSG00000026471 - Ensembl, May 2017
- ↑ "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ↑ "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- 1 2 3 4 5 6 7 8 Krovi SH, Gapin L (August 2016). "Structure and function of the non-classical major histocompatibility complex molecule MR1". Immunogenetics. 68 (8): 549–559. doi:10.1007/s00251-016-0939-5. PMC 5091073. PMID 27448212.
- ↑ McWilliam HE, Villadangos JA (September 2017). "How MR1 Presents a Pathogen Metabolic Signature to Mucosal-Associated Invariant T (MAIT) Cells". Trends in Immunology. 38 (9): 679–689. doi:10.1016/j.it.2017.06.005. PMID 28688841.
- 1 2 3 Corbett AJ, Eckle SB, Birkinshaw RW, Liu L, Patel O, Mahony J, et al. (May 2014). "T-cell activation by transitory neo-antigens derived from distinct microbial pathways". Nature. 509 (7500): 361–365. Bibcode:2014Natur.509..361C. doi:10.1038/nature13160. PMID 24695216. S2CID 4401282.
- 1 2 3 Kjer-Nielsen L, Patel O, Corbett AJ, Le Nours J, Meehan B, Liu L, et al. (November 2012). "MR1 presents microbial vitamin B metabolites to MAIT cells" (PDF). Nature. 491 (7426): 717–723. Bibcode:2012Natur.491..717K. doi:10.1038/nature11605. PMID 23051753. S2CID 4419703.
- ↑ "UniProt, Q95460".
- ↑ Karamooz, Elham; Harriff, Melanie J.; Lewinsohn, David M. (December 2018). "MR1-dependent antigen presentation". Seminars in Cell & Developmental Biology. 84: 58–64. doi:10.1016/j.semcdb.2017.11.028. ISSN 1096-3634. PMC 7061520. PMID 30449535.
- ↑ Jeffery, Hannah C.; van Wilgenburg, Bonnie; Kurioka, Ayako; Parekh, Krishan; Stirling, Kathryn; Roberts, Sheree; Dutton, Emma E.; Hunter, Stuart; Geh, Daniel; Braitch, Manjit K.; Rajanayagam, Jeremy (May 2016). "Biliary epithelium and liver B cells exposed to bacteria activate intrahepatic MAIT cells through MR1". Journal of Hepatology. 64 (5): 1118–1127. doi:10.1016/j.jhep.2015.12.017. ISSN 1600-0641. PMC 4822535. PMID 26743076.
- ↑ Lett, Martin J.; Mehta, Hema; Keogh, Adrian; Jaeger, Tina; Jacquet, Maxime; Powell, Kate; Meier, Marie-Anne; Fofana, Isabel; Melhem, Hassan; Vosbeck, Jürg; Cathomas, Gieri (2022-01-20). "Stimulatory MAIT cell antigens reach the circulation and are efficiently metabolised and presented by human liver cells". Gut. 71 (12): gutjnl–2021–324478. doi:10.1136/gutjnl-2021-324478. ISSN 1468-3288. PMC 9664123. PMID 35058274. S2CID 246081888.
- 1 2 Mori L, Lepore M, De Libero G (May 2016). "The Immunology of CD1- and MR1-Restricted T Cells". Annual Review of Immunology. 34 (1): 479–510. doi:10.1146/annurev-immunol-032414-112008. PMID 26927205.
- 1 2 McWilliam HE, Mak JY, Awad W, Zorkau M, Cruz-Gomez S, Lim HJ, et al. (October 2020). "Endoplasmic reticulum chaperones stabilize ligand-receptive MR1 molecules for efficient presentation of metabolite antigens". Proceedings of the National Academy of Sciences of the United States of America. 117 (40): 24974–24985. Bibcode:2020PNAS..11724974M. doi:10.1073/pnas.2011260117. PMC 7547156. PMID 32958637.
- ↑ McWilliam HE, Eckle SB, Theodossis A, Liu L, Chen Z, Wubben JM, et al. (May 2016). "The intracellular pathway for the presentation of vitamin B-related antigens by the antigen-presenting molecule MR1" (PDF). Nature Immunology. 17 (5): 531–537. doi:10.1038/ni.3416. PMID 27043408. S2CID 23473219.
- ↑ Keller AN, Eckle SB, Xu W, Liu L, Hughes VA, Mak JY, et al. (April 2017). "Drugs and drug-like molecules can modulate the function of mucosal-associated invariant T cells" (PDF). Nature Immunology. 18 (4): 402–411. doi:10.1038/ni.3679. hdl:11343/290500. PMID 28166217. S2CID 3546821.
- ↑ Mak JY, Liu L, Fairlie DP (September 2021). "Chemical Modulators of Mucosal Associated Invariant T Cells". Accounts of Chemical Research. 54 (17): 3462–3475. doi:10.1021/acs.accounts.1c00359. PMC 8989627. PMID 34415738.
- ↑ Veerapen N, Hobrath J, Besra AK, Besra GS (January 2021). "Chemical insights into the search for MAIT cells activators". Molecular Immunology. 129: 114–120. doi:10.1016/j.molimm.2020.11.017. PMID 33293098. S2CID 228078338.
- ↑ Mak JY, Xu W, Reid RC, Corbett AJ, Meehan BS, Wang H, et al. (March 2017). "Stabilizing short-lived Schiff base derivatives of 5-aminouracils that activate mucosal-associated invariant T cells". Nature Communications. 8 (1): 14599. Bibcode:2017NatCo...814599M. doi:10.1038/ncomms14599. PMC 5344979. PMID 28272391.
- ↑ Karamooz E, Harriff MJ, Lewinsohn DM (December 2018). "MR1-dependent antigen presentation". Seminars in Cell & Developmental Biology. 84: 58–64. doi:10.1016/j.semcdb.2017.11.028. PMC 7061520. PMID 30449535.
- 1 2 McWilliam HE, Villadangos JA (June 2020). "MR1: a multi-faceted metabolite sensor for T cell activation". Current Opinion in Immunology (review). 64: 124–129. doi:10.1016/j.coi.2020.05.006. hdl:11343/267503. PMID 32604057. S2CID 220282936.
- ↑ Crowther MD, Dolton G, Legut M, Caillaud ME, Lloyd A, Attaf M, et al. (February 2020). "Genome-wide CRISPR-Cas9 screening reveals ubiquitous T cell cancer targeting via the monomorphic MHC class I-related protein MR1". Nature Immunology (primary). 21 (2): 178–185. doi:10.1038/s41590-019-0578-8. PMC 6983325. PMID 31959982.
- ↑ Vacchini A, Chancellor A, Spagnuolo J, Mori L, De Libero G (2020-04-28). "MR1-Restricted T Cells Are Unprecedented Cancer Fighters". Frontiers in Immunology (review). 11: 751. doi:10.3389/fimmu.2020.00751. PMC 7198878. PMID 32411144.
- ↑ Yan J, Allen S, McDonald E, Das I, Mak JY, Liu L, et al. (January 2020). "MAIT Cells Promote Tumor Initiation, Growth, and Metastases via Tumor MR1". Cancer Discovery (primary). 10 (1): 124–141. doi:10.1158/2159-8290.CD-19-0569. PMID 31826876. S2CID 209329710.
- ↑ Li, Wei-Zheng; Stirling, Kyle; Yang, Jun-Jie; Zhang, Lei (2020-07-15). "Gut microbiota and diabetes: From correlation to causality and mechanism". World Journal of Diabetes. 11 (7): 293–308. doi:10.4239/wjd.v11.i7.293. ISSN 1948-9358. PMC 7415231. PMID 32843932.
- 1 2 Toubal A, Kiaf B, Beaudoin L, Cagninacci L, Rhimi M, Fruchet B, et al. (July 2020). "Mucosal-associated invariant T cells promote inflammation and intestinal dysbiosis leading to metabolic dysfunction during obesity". Nature Communications (primary). 11 (1): 3755. Bibcode:2020NatCo..11.3755T. doi:10.1038/s41467-020-17307-0. PMC 7381641. PMID 32709874.
- ↑ Rouxel O, Da Silva J, Beaudoin L, Nel I, Tard C, Cagninacci L, et al. (December 2017). "Cytotoxic and regulatory roles of mucosal-associated invariant T cells in type 1 diabetes". Nature Immunology (primary). 18 (12): 1321–1331. doi:10.1038/ni.3854. PMC 6025738. PMID 28991267.
- ↑ Haga K, Chiba A, Shibuya T, Osada T, Ishikawa D, Kodani T, et al. (May 2016). "MAIT cells are activated and accumulated in the inflamed mucosa of ulcerative colitis" (PDF). Journal of Gastroenterology and Hepatology (primary). 31 (5): 965–972. doi:10.1111/jgh.13242. PMID 26590105. S2CID 38327157.
- ↑ Yasutomi Y, Chiba A, Haga K, Murayama G, Makiyama A, Kuga T, et al. (2022-01-01). "Activated Mucosal-associated Invariant T Cells Have a Pathogenic Role in a Murine Model of Inflammatory Bowel Disease". Cellular and Molecular Gastroenterology and Hepatology (primary). 13 (1): 81–93. doi:10.1016/j.jcmgh.2021.08.018. PMC 8593615. PMID 34461283.
- 1 2 Mortier C, Govindarajan S, Venken K, Elewaut D (2018). "It Takes "Guts" to Cause Joint Inflammation: Role of Innate-Like T Cells". Frontiers in Immunology (review). 9: 1489. doi:10.3389/fimmu.2018.01489. PMC 6033969. PMID 30008717.
- ↑ Argollo M, Gilardi D, Peyrin-Biroulet C, Chabot JF, Peyrin-Biroulet L, Danese S (August 2019). "Comorbidities in inflammatory bowel disease: a call for action". The Lancet. Gastroenterology & Hepatology (review). 4 (8): 643–654. doi:10.1016/S2468-1253(19)30173-6. PMID 31171484. S2CID 174814922.
- ↑ Koppejan, Hester; Jansen, Diahann T. S. L.; Hameetman, Marjolijn; Thomas, Ranjeny; Toes, Rene E. M.; van Gaalen, Floris A. (2019-01-05). "Altered composition and phenotype of mucosal-associated invariant T cells in early untreated rheumatoid arthritis". Arthritis Research & Therapy. 21 (1): 3. doi:10.1186/s13075-018-1799-1. ISSN 1478-6362. PMC 6321723. PMID 30611306.
- ↑ Hayashi, Eri; Chiba, Asako; Tada, Kurisu; Haga, Keiichi; Kitagaichi, Mie; Nakajima, Shihoko; Kusaoi, Makio; Sekiya, Fumio; Ogasawara, Michihiro; Yamaji, Ken; Tamura, Naoto (2016-09-01). "Involvement of Mucosal-associated Invariant T cells in Ankylosing Spondylitis". The Journal of Rheumatology. 43 (9): 1695–1703. doi:10.3899/jrheum.151133. ISSN 0315-162X. PMID 27370879. S2CID 40101757.
- ↑ Gracey, Eric; Qaiyum, Zoya; Almaghlouth, Ibrahim; Lawson, Daeria; Karki, Susan; Avvaru, Naga; Zhang, Zhenbo; Yao, Yuchen; Ranganathan, Vidya; Baglaenko, Yuriy; Inman, Robert D. (2016-12-01). "IL-7 primes IL-17 in mucosal-associated invariant T (MAIT) cells, which contribute to the Th17-axis in ankylosing spondylitis". Annals of the Rheumatic Diseases. 75 (12): 2124–2132. doi:10.1136/annrheumdis-2015-208902. ISSN 0003-4967. PMID 27165176. S2CID 206851982.
Further reading
- Maruyama K, Sugano S (January 1994). "Oligo-capping: a simple method to replace the cap structure of eukaryotic mRNAs with oligoribonucleotides". Gene. 138 (1–2): 171–174. doi:10.1016/0378-1119(94)90802-8. PMID 8125298.
- Yamaguchi H, Hirai M, Kurosawa Y, Hashimoto K (September 1997). "A highly conserved major histocompatibility complex class I-related gene in mammals". Biochemical and Biophysical Research Communications. 238 (3): 697–702. doi:10.1006/bbrc.1997.7379. PMID 9325151.
- Suzuki Y, Yoshitomo-Nakagawa K, Maruyama K, Suyama A, Sugano S (October 1997). "Construction and characterization of a full length-enriched and a 5'-end-enriched cDNA library". Gene. 200 (1–2): 149–156. doi:10.1016/S0378-1119(97)00411-3. PMID 9373149.
- Riegert P, Wanner V, Bahram S (October 1998). "Genomics, isoforms, expression, and phylogeny of the MHC class I-related MR1 gene". Journal of Immunology. 161 (8): 4066–4077. doi:10.4049/jimmunol.161.8.4066. PMID 9780177. S2CID 23819173.
- Parra-Cuadrado JF, Navarro P, Mirones I, Setién F, Oteo M, Martínez-Naves E (August 2000). "A study on the polymorphism of human MHC class I-related MR1 gene and identification of an MR1-like pseudogene". Tissue Antigens. 56 (2): 170–172. doi:10.1034/j.1399-0039.2000.560211.x. PMID 11019920.
- Miley MJ, Truscott SM, Yu YY, Gilfillan S, Fremont DH, Hansen TH, Lybarger L (June 2003). "Biochemical features of the MHC-related protein 1 consistent with an immunological function". Journal of Immunology. 170 (12): 6090–6098. doi:10.4049/jimmunol.170.12.6090. PMID 12794138.
- Hempelmann A, Kumar S, Muralitharan S, Sander T (July 2006). "Myofibrillogenesis regulator 1 gene (MR-1) mutation in an Omani family with paroxysmal nonkinesigenic dyskinesia". Neuroscience Letters. 402 (1–2): 118–120. doi:10.1016/j.neulet.2006.03.048. PMID 16632198. S2CID 22214392.