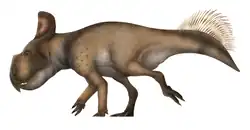| Nasutoceratops Temporal range: Campanian, | |
|---|---|
 | |
| Reconstructed skull at Arizona Museum of Natural History | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Clade: | †Ornithischia |
| Suborder: | †Ceratopsia |
| Family: | †Ceratopsidae |
| Subfamily: | †Centrosaurinae |
| Tribe: | †Nasutoceratopsini |
| Genus: | †Nasutoceratops Sampson et al., 2013 |
| Type species | |
| †Nasutoceratops titusi Sampson et al., 2013 | |
Nasutoceratops is an extinct genus of ceratopsian dinosaur. It is a basal centrosaurine which lived during the Late Cretaceous Period (late Campanian, about 76.0-75.5 Ma). Fossils have been found in southern Utah, United States. Nasutoceratops was a large, ground-dwelling, quadrupedal herbivore with a short snout and unique rounded horns above its eyes that have been likened to those of modern cattle. Extending almost to the tip of its snout, these horns are the longest of all the members of the centrosaurine subfamily. The presence of pneumatic elements in the nasal bones of Nasutoceratops are a unique trait and are unknown in any other ceratopsid.
Discovery and naming

Since 2000, the Natural History Museum of Utah (UMNH) and the Bureau of Land Management have been conducting paleontological surveys of the Kaiparowits Formation at the Grand Staircase–Escalante National Monument (GSENM) in southern Utah. This national monument was established in 1996 in part for the preservation and study of its fossils, and the surveys there have yielded a wide array of unique dinosaur fossils. Field crews from other institutions have also participated, and the collaborative effort has been called the Kaiparowits Basin Project.[1][2] Among the discoveries made were three new ceratopsian (horned dinosaur) taxa, one of which was identified from UMNH Locality VP 940 discovered by then graduate student and technician Eric K. Lund during the 2006 field season.[2][3]
Prior to this project, the only ceratopsian remains found at the formation were uninformative, isolated teeth.[4] Excavated fossils were transported to the UMNH, where the blocks were prepared by volunteers with pneumatic air scribes and needles and subsequently reassembled; it took a few years for the team to assemble the skull of this dinosaur.[1][5][6] It was preliminarily referred to as "Kaiparowits new taxon C" and identified as a centrosaurine ceratopsid (the first centrosaurine known from the formation) in 2010, and as "Kaiparowits centrosaurine A" in 2013. Three specimens of this taxon were collected; UMNH VP 16800 in 2006, with UMNH VP 19469 and UMNH VP 19466 collected in subsequent years.[2][3][7]
The paleontologists Scott D. Sampson, Lund, Mark A. Loewen, Andrew A. Farke and Katherine E. Clayton briefly described and scientifically named the new genus and species Nasutoceratops titusi in 2013, with specimen UMNH VP 16800 as the holotype (on which the scientific name is based). The generic name is derived from the Latin word nasutus meaning "large-nosed", and ceratops, which means "horned-face" in Latinized Greek. The specific name titusi is an eponym that honors the American paleotologist Alan L. Titus for his important efforts in recovering fossils from the GSENM.[8][4] Lund had informally used the spelling Nasutuceratops for this dinosaur in his 2010 thesis.[9] In 2016, Lund, Sampson, and Loewen published a more detailed description of the preserved fossil material.[4]



The holotype specimen UMNH VP 16800 consists of a partial, associated, and nearly complete skull of a subadult that preserves most of the skull roof, which was collected with an articulated and almost complete left forelimb, an associated yet very fragmentary right hindlimb, much of the pectoral girdle, an almost complete syncervical vertebra, and three associated but fragmentary dorsal vertebrae, as well as skin impressions associated with the left forelimb. Two specimens were assigned due to shared features with the holotype: specimen UMNH VP 19466, a disarticulated adult skull consisting of the partial right and left premaxillae, a right maxilla and right nasal, and specimen UMNH VP 19469, an isolated squamosal bone of a subadult. Taken together, these specimens represent about 80% of the skull and about 10% of the postcranial skeleton.[8][4]
In the UMNH press release accompanying the description of Nasutuceratops, the large nose of the dinosaur was emphasized, with Sampson calling it a "jumbo-sized schnoz".[7] This was reflected by news outlets, with one article titled "Paleontologists Discover, Mock, New Dinosaur Species", and another including humorous poems about the dinosaur by columnist Alexandra Petri, such as: "Higgledy piggledy, Nasutoceratops, Long-nosed horned just-unearthed dino du jour, Probably used its horns, For showing dominance, During its courtship (although we’re not sure.)"[10][11][12][13][6] Nasutuceratops was featured in the 2019 Jurassic World short film Battle at Big Rock and the 2022 film Jurassic World Dominion, in what an UMNH article called a "pivotal role".[14][15]
In 2017, the US government announced plans to shrink the GSENM (to little over half its size) and Bears Ears monuments to enable coal mining and other energy development on the land, which was the largest reduction of US national monuments in history.[16][17] Sampson, who had overseen much of the early research at the former monument, expressed fear that such a move would threaten further discoveries. Media outlets stressed the importance of the area's fossil discoveries—including more than 25 new taxa—with some noting the discovery of Nasutoceratops.[17][18][19] The US government was subsequently sued by a group of scientists, environmentalists, and Native Americans; in 2021, the monument was restored to its former extent by the new administration.[16][20][21]
Description

The holotype skull is approximately 1.5 metres (4.9 ft) in length. Its body length has been estimated at 4.5 metres (14.8 ft), its weight at 1.5 tonnes (1.7 short tons).[22] Nasutoceratops has several unique derived traits or autapomorphies. The part of the snout around the nostril is strongly developed, representing about three quarters of the skull length in front of the eye sockets. The rear of each nasal bone is hollowed out by a large internal cavity. The contact surface between the maxilla and the praemaxilla is exceptionally large. The maxilla also has a large internal flange contacting the praemaxilla via two horizontal facets. The brow horns at their bases are pointed forwards and outwards, then curve inwards, and ultimately twist their points upwards.[8]

Nasutoceratops also shows a unique combination of traits that are in themselves not unique. The horn on the nose is low and narrow, with an elongated base. The squamosal has a high ridge on its upper surface, running from the direction of the eye socket towards the squamosal rim. The skull frill is more or less circular with its widest point at the middle edge. The osteoderms on the frill edge, the epiparietals and episquamosals, do not have the form of spikes but are shaped like simple low crescents. The rear frill edge is not notched but instead has an epiparietal on the midline.[8]
The snout of Nasutoceratops was short and high; its nasal bones exhibit internal cavities that the authors consider to represent pneumatic excavations, invading the bone from the nasal cavity. This is noteworthy because pneumatic nasals are unknown in any other ceratopsid, which supports that this feature represents a unique derived trait of this genus. Nasutoceratops had as many as 29 tooth positions in the maxilla, each occupied by several stacked teeth. The skull roof between the eye sockets is vaulted and is markedly higher than the snout region. The curved horizontally projecting brow horn arrangement was likened to that of modern cattle by paleontologist David Hone.[23] The brow horns span approximately 40% of the total length of the skull, almost reaching the level of the snout tip, and with a bone core length of up to 457 millimetres are the longest known of any centrosaurine, both in absolute and relative terms.[9]

The epijugal (cheek horn) has a length of 85 millimetres (3.3 in), also the largest known among centrosaurines. The skull frill is moderately long, and pierced by a large kidney-shaped parietal fenestra at each side. Apart from the midline epiparietal, there are seven epiparietals at each side, and about four to five episquamosals.[9] In the forelimb, the ulna is exceptionally robust with a large olecranon process. Of the three patches with skin impressions found near the left shoulder, one shows a pattern of large, eight to eleven millimetres wide, hexagonal scales surrounded by smaller triangular scales.[9][24]
Classification



Nasutoceratops was assigned to the Centrosaurinae in 2013, in a relatively basal position. A phylogenetic analysis performed by Sampson et al. (2013) found Nasutoceratops to be the sister taxon of Avaceratops. According to this study, the existence of Nasutoceratops would support the hypothesis of faunal separation between the north and south of Laramidia. Its clade would differ from the northern centrosaurines in the retention of long brow horns and a short nose horn, combined with developing, convergent with the Chasmosaurinae, low epiparietals.[8] In 2016, this clade was named Nasutoceratopsini; it contains Nasutoceratops as well as ANSP 15800 (the holotype of Avaceratops), MOR 692 (previously treated as an adult specimen of Avaceratops), the newly described CMN 8804 from the Oldman Formation, and another undescribed ceratopsian found in Malta, Montana.[25] The cladogram presented below follows a phylogenetic analysis of the Centrosaurinae by Chiba et al. (2017), which included a systematic re-evaluation of Medusaceratops lokii:[26][27][28]
| Centrosaurinae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiology

In 2016, Lund and colleagues stated that the functional adaptations associated with the distinct cranial features of Nasutoceratops are unknown. They suggested that the very short and deep front part of the skull in front of the eyes may have been connected to a change toward more derived masticatory functions in basal ceratopsians. This morphology increased the mechanical advantage during mastication by moving the beak closer to the joint of the lower jaw. The narial region of Nasutoceratops is deep mainly due to the premaxilla and maxilla having steeply rising, enlarged contact surfaces, but the function of this is unknown. because the contact surfaces are steeply inclined and more robust than those o more derived relatives, this feature may be connected with absorbing larger bite forces. The pneumaticity in the nasal may have derived from a source in front of the nasal. Pneumaticity in this region has been associated with a variety of functions in vertebrates such as moisture exchange, shock absorption, vocal resonance, and weight reduction, but the function in Nasutoceratops is unclear.[4] Sampson stated in a 2013 press release that the large snout probably did not have anything to do with a heightened sense of smell, since olfactory sensors are located further back in the head, closer to the brain.[7]
In a 2017 Master's thesis, paleontologist Nicole Marie Ridgwell described two coprolites (fossilized dung) from the Kaiparowits Formation which, due to their size, may have been produced by a member of one of three herbivorous dinosaur groups known from the formation: ceratopsians (including Nasutoceratops), hadrosaurs, or ankylosaurs (rarest of the three groups). The coprolites contained fragments of angiosperm wood (which indicates a diet of woody browse); though there was previously little evidence of dinosaurs consuming angiosperms, these coprolites showed that dinosaurs adapted to feeding on them (they only became common in the Early Cretaceous, diversifying in the Late Cretaceous). The coprolites also contained traces of mollusc shell, arthropod cuticle, and lizard bone that may have been ingested along with the plant material. They were found near other herbivore coprolites that contained conifer wood. Ridgwell pointed out that the dental anatomies of ceratopsians and hadrosaurs (with dental batteries comprising continuously replaced teeth) were adapted to process large quantities of fibrous plants. The different diets represented by the coprolites may indicate niche partitioning among the herbivores of the Kaiparowits Formation ecosystem, or that there was seasonal variation in diet.[29]
Function of skull ornamentation

Sampson and colleagues stated in 2013 that while various hypotheses about the function of ceratopsid ornamentation have been proposed, the consensus at the time was use in intraspecific signalling and intraspecific combat, with the debate focusing on either species recognition (driven by natural selection) or mate competition (driven by sexual selection). They pointed out that either way, the evolutionary changes in the two centrosaurine clades were concentrated in different parts of the skull, with the clade that included Avaceratops and Nasutoceratops reducing frill ornamentation but elaborating the supraorbital horns, and the clade that indluded Spinops and Pachyrhinosaurus clade reducing supraorbital horns but hypertrophying nasal horn and frill orientation, which distributed their distinct signalling structures all across the skull roof.[9] Loewen stated in the 2013 press release that the horns were probably used as visual signs of dominance, and as weapons against rivals when that was not enough.[7]
In 2016, Lund and colleagues suggested that if the mate competition hypothesis applied to the very long and robust supraorbital horns of Nasutoceratops, their orientation towards the front and sides and torsional twist may have enabled interlocking of horns with opponents of the same species, as seen in many modern bovids. They noted that the paleontologist Andrew A. Farke had earlier studied the function of ceratopsid supraorbital horns by using scale models, and found three plausible horn locking positions for Triceratops that could also apply to Nasutoceratops; "single horn contact", "full horn locking", and "oblique horn locking". Pathologies in the skulls of Triceratops and Centrosaurus, with those in the former concluded to be consistent with trauma resulting from antagonistic behaviour, but those of the latter not as conclusive, and Lund and colleagues found that such hypothesis could not be ruled out for Nasutoceratops.[4]
Paleoenvironment
.jpg.webp)

Nasutoceratops is known from the Kaiparowits Formation of Utah, which dates to the late Campanian age of the Late Cretaceous epoch, and stratigraphically occurs in the formation's middle unit, in sediments dating to 75.51-75.97 Ma million years ago. The formation was deposited in the southern part of a basin (the Western Interior Basin) on the eastern margin of a landmass known as Laramidia (an island continent consisting of what is now western North America) within 100 km (62 mi) of the Western Interior Seaway, a shallow sea in the center of North America that divided the continent (the eastern landmass is known as Appalachia).[9][30][31] The basin was broad, flat, crescent-shaped, and bounded by mountains on all sides except the Western Interior Seaway at the east.[32] The formation represents an alluvial to coastal plain setting that was wet, humid, and dominated by large, deep channels with stable banks and perennial wetland swamps, ponds, and lakes. Rivers flowed generally west across the plains and drained into the Western Interior Seaway; the Gulf Coast region of the United States has been proposed as a good modern analogue (such as the current day swamplands of Louisiana). The formation preserves a diverse and abundant range of fossils, including continental and aquatic animals, plants, and palynomorphs (organic microfossils).[33][5]

Other ornithischian dinosaurs from the Kaiparowits Formation include ceratopsians such as the chasmosaurines Utahceratops and Kosmoceratops (and possibly a second yet unnamed centrosaurine), indeterminate pachycephalosaurs, the ankylosaurid Akainacephalus, an indeterminate nodosaurid, the hadrosaurs Gryposaurus and Parasaurolophus, and an indeterminate, basal neornithischian. Theropods include the tyrannosaurid Teratophoneus, the oviraptorosaur Hagryphus, an unnamed ornithomimid, the troodontid Talos, indeterminate dromaeosaurids, and the bird Avisaurus. Other vertebrates include crocodiles (such as Deinosuchus and Brachychampsa), turtles (such as Adocus and Basilemys), pterosaurs, lizards, snakes, amphibians, mammals, and fishes.[32][34][35] The two most common groups of large vertebrates in the formation are hadrosaurs and ceratopsians (the latter representing about 14 percent of associated vertebrate fossils), which may either indicate their abundance in the Kaiparowits fauna or reflect preservation bias (a type of sampling bias) due to these groups also having the most robust skeletal elements.[3] Eggs from dinosaurs, crocodiles, and turtles have also been found.[36] The swamps and wetlands were dominated by up to 30 m (98 ft) cypress trees, ferns, and aquatic plants including giant duckweed, water lettuce, and other floating angiosperms. Better-drained areas were dominated by forests of up to 10–20 m (33–66 ft) dicot trees and occasional palms, with an understory including ferns. Well-drained areas further away from wet areas were dominated by conifers up to 30 m (98 ft), with an understory comprising cycads, small dicot trees or bushes, and possibly ferns.[32]
See also
References
- 1 2 Sampson, S. D.; Loewen, M. A.; Farke, A. A.; Roberts, E. M.; Forster, C. A.; Smith, J. A.; Titus, A. L.; Stepanova, A. (2010). "New horned dinosaurs from Utah provide evidence for intracontinental dinosaur endemism". PLOS ONE. 5 (9): e12292. Bibcode:2010PLoSO...512292S. doi:10.1371/journal.pone.0012292. PMC 2929175. PMID 20877459.
- 1 2 3 Loewen, M.; Farke, A. A.; Sampson, S. D.; Getty, M. A.; Lund, E. K.; O’Connor, P. M. (2013). "Ceratopsid dinosaurs from the Grand Staircase of Southern Utah". In Titus, A.; Loewen, M. (eds.). At the Top of the Grand Staircase: The Late Cretaceous of Southern Utah. Bloomington: Indiana University Press. pp. 488–503. ISBN 978-0-253-00883-1.
- 1 2 3 Getty, M. A.; Loewen, M. A.; Roberts, E.; Titus, A. L.; Sampson, S. D. (2010), "Taphonomy of horned dinosaurs (Ornithischia: Ceratopsidae) from the late Campanian Kaiparowits Formation, Grand Staircase – Escalante National Monument, Utah", in Ryan, M. J.; Chinnery-Allgeier, B. J.; Eberth, D. A. (eds.), New Perspectives on Horned Dinosaurs: The Royal Tyrrell Museum Ceratopsian Symposium, Bloomington: Indiana University Press, pp. 478–494, ISBN 978-0253353580
- 1 2 3 4 5 6 Lund, Eric K.; Sampson, Scott D.; Loewen, Mark A. (2016). "Nasutoceratops titusi (Ornithischia, Ceratopsidae), a basal centrosaurine ceratopsid from the Kaiparowits Formation, southern Utah". Journal of Vertebrate Paleontology. 36 (2): e1054936. doi:10.1080/02724634.2015.1054936.
- 1 2 Sampson, S. D. (2012). "Dinosaurs of the lost continent". Scientific American. 306 (3): 40–47. Bibcode:2012SciAm.306c..40S. doi:10.1038/scientificamerican0312-40. ISSN 0036-8733. JSTOR 26014255. PMID 22375321.
- 1 2 "Dinosaur gets named for his really big nose". The Denver Post. 2013. Retrieved 9 January 2024.
- 1 2 3 4 Carpenter, Patti (8 October 2013). "Big-Nosed, Long-Horned Dinosaur Discovered in Utah". Natural History Museum of Utah. Retrieved 9 January 2024.
- 1 2 3 4 5 Sampson, S. D.; Lund, E. K.; Loewen, M. A.; Farke, A. A.; Clayton, K. E. (2013). "A remarkable short-snouted horned dinosaur from the Late Cretaceous (late Campanian) of southern Laramidia". Proceedings of the Royal Society B: Biological Sciences. 280 (1766): 20131186. doi:10.1098/rspb.2013.1186. PMC 3730592. PMID 23864598. full text
- 1 2 3 4 5 6 Eric Karl Lund (2010). Nasutuceratops titusi, a new basal centrosaurine dinosaur (Ornithischia: Ceratopsidae) from the upper cretaceous Kaiparowits Formation, Southern Utah. Department of Geology and Geophysics University of Utah (Thesis). pp. 172 pp. Archived from the original on 2015-04-02.
- ↑ Ohlheiser, Abby (18 July 2013). "Paleontologists Discover, Mock, New Dinosaur Species". The Atlantic. Retrieved 9 January 2024.
- ↑ Petri, Alexandra (2 December 2021). "Three-horned poems for the new dinosaur, Nasutoceratops, relative of the triceratops". Washington Post. Retrieved 9 January 2024.
- ↑ Memmott, Mark (2013). "Newly Discovered Dinosaur Sure Had One 'Supersize Schnoz'". NPR. Retrieved 9 January 2024.
- ↑ Dell'Amore, Christine (4 November 2013). "New Big-Nosed Horned Dinosaur Found in Utah". National Geographic News. Retrieved 9 January 2024.
- ↑ Whalen, Andrew (16 September 2019). "All 7 Dinosaurs in 'Battle at Big Rock,' Including Nasutoceratops". Newsweek. Retrieved 9 January 2024.
- ↑ Irmis, Randall (21 June 2022). "NHMU Dinosaur Stars in Jurassic World Dominion | Natural History Museum of Utah". nhmu.utah.edu. Retrieved 9 January 2024.
- 1 2 Gonzales, R.; Siegler, K.; Dwyer, C. (2017). "Trump orders largest national monument reduction In U.S. history". NPR.org. Archived from the original on July 5, 2019. Retrieved June 25, 2019.
- 1 2 Finnegan, M. (2017). "Remarkable dinosaur discoveries under threat with Trump plan to shrink national monument in Utah, scientists say". Los Angeles Times. Archived from the original on June 25, 2019. Retrieved June 24, 2019.
- ↑ Worby, Rebecca (3 November 2017). "What could be lost in a push for mining in monuments". www.hcn.org. Retrieved 28 December 2023.
- ↑ Panciroli, E. (2017). "Why are palaeontologists suing Trump?". The Guardian. Archived from the original on May 31, 2019. Retrieved June 25, 2019.
- ↑ Wei-Haas, M. (2017). "What shrinking fossil-rich national monuments means for science". Smithsonian Magazine. Archived from the original on June 24, 2019. Retrieved June 24, 2019.
- ↑ "The Wilderness Society et al.v. Trump et al. (Grand Staircase–Escalante)". NRDC. April 10, 2020. Retrieved April 19, 2020.
- ↑ Paul, G.S., 2016, The Princeton Field Guide to Dinosaurs 2nd edition, Princeton University Press p. 287
- ↑ Hone, Dave (2013-07-17). "Newly discovered dinosaur Nasutoceratops had cow-like horns". guardian.co.uk. London, UK: Guardian. Retrieved 2013-07-17.
- ↑ Bell, Phil R.; Hendrickx, Christophe; Pittman, Michael; Kaye, Thomas G.; Mayr, Gerald (2022). "The exquisitely preserved integument of Psittacosaurus and the scaly skin of ceratopsian dinosaurs". Communications Biology. 5 (1). doi:10.1038/s42003-022-03749-3. PMC 9374759.
- ↑ Ryan, M.J.; Holmes, R.; Mallon, J.; Loewen, M.; Evans, D.C. (2017). "A basal ceratopsid (Centrosaurinae: Nasutoceratopsini) from the Oldman Formation (Campanian) of Alberta, Canada". Canadian Journal of Earth Sciences. 54 (1): 1–14. Bibcode:2017CaJES..54....1R. doi:10.1139/cjes-2016-0110.
- ↑ Chiba, Kentaro; Ryan, Michael J.; Fanti, Federico; Loewen, Mark A.; Evans, David C. (2018). "New material and systematic re-evaluation of Medusaceratops lokii (Dinosauria, Ceratopsidae) from the Judith River Formation (Campanian, Montana)". Journal of Paleontology. 92 (2): 272–288. Bibcode:2018JPal...92..272C. doi:10.1017/jpa.2017.62. S2CID 134031275.
- ↑ Lund, Eric K.; O’Connor, Patrick M.; Loewen, Mark A.; Jinnah, Zubair A. (2016). "A new centrosaurine ceratopsid, Machairoceratops cronusi gen et sp. nov., from the Upper Sand Member of the Wahweap Formation (Middle Campanian), Southern Utah". PLOS ONE. 11 (5): e0154403. Bibcode:2016PLoSO..1154403L. doi:10.1371/journal.pone.0154403.
- ↑ Rivera-Sylva, Héctor E.; Hedrick, Brandon P.; Dodson, Peter (2016). "A Centrosaurine (Dinosauria: Ceratopsia) from the Aguja Formation (Late Campanian) of Northern Coahuila, Mexico". PLOS ONE. 11 (4): e0150529. Bibcode:2016PLoSO..1150529R. doi:10.1371/journal.pone.0150529. PMID 27073969.
- ↑ Ridgwell, N. M. (2017). Description of Kaiparowits coprolites that provide rare direct evidence of angiosperm consumption by dinosaurs. Museum and Field Studies Graduate Theses & Dissertations (Thesis). Archived from the original on June 30, 2019. Retrieved June 30, 2019.
- ↑ Roberts, E. M.; Deino, A. L.; Chan, M. A. (2005). "40Ar/39Ar age of the Kaiparowits Formation, southern Utah, and correlation of contemporaneous Campanian strata and vertebrate faunas along the margin of the Western Interior Basin". Cretaceous Research. 26 (2): 307–318. Bibcode:2005CrRes..26..307R. doi:10.1016/j.cretres.2005.01.002.
- ↑ Fowler, D. W.; Wong, William O. (2017). "Revised geochronology, correlation, and dinosaur stratigraphic ranges of the Santonian-Maastrichtian (Late Cretaceous) formations of the Western Interior of North America". PLOS ONE. 12 (11): e0188426. Bibcode:2017PLoSO..1288426F. doi:10.1371/journal.pone.0188426. PMC 5699823. PMID 29166406.
- 1 2 3 Sampson, S. D.; Loewen, M. A.; Roberts, E. M.; Getty, M. A. (2013). "A new macrovertebrate assemblage from the Late Cretaceous (Campanian) of Southern Utah". In Titus, A.; Loewen, M. (eds.). At the Top of the Grand Staircase: The Late Cretaceous of Southern Utah. Bloomington: Indiana University Press. pp. 599–622. ISBN 978-0-253-00883-1.
- ↑ Roberts, E. M.; Sampson, S. D.; Deino, A. L.; Bowring, S. A.; Buchwaldt, S. (2013). "The Kaiparowits Formation: a remarkable record of Late Cretaceous terrestrial environments, ecosystems, and evolution in Western North America". In Titus, A.; Loewen, M. (eds.). At the Top of the Grand Staircase: The Late Cretaceous of Southern Utah. Bloomington: Indiana University Press. pp. 85–106. ISBN 978-0-253-00883-1.
- ↑ Titus, A. L.; Eaton, J. G.; Sertich, J. (2016). "Late Cretaceous stratigraphy and vertebrate faunas of the Markagunt, Paunsaugunt, and Kaiparowits plateaus, southern Utah". Geology of the Intermountain West. 3: 229–291. doi:10.31711/giw.v3i0.10.
- ↑ Wiersma, J. P.; Irmis, R. B. (2018). "A new southern Laramidian ankylosaurid, Akainacephalus johnsoni gen. et sp. nov., from the upper Campanian Kaiparowits Formation of southern Utah, USA". PeerJ. 6: 76. doi:10.7717/peerj.5016. PMC 6063217. PMID 30065856.
- ↑ Oser, S. E. (2018). Campanian ooassemblages within the Western Interior Basin: eggshell from the Upper Cretaceous Kaiparowits Formation of Grand Staircase–Escalante National Monument, UT (MSc thesis). Boulder: Department of Museum and Field Studies, University of Colorado. S2CID 134887908.
External links
- Meet Nasutoceratops: Big-Nose Horned Face - 11 minute NPR audio interview with describer Scott D. Sampson
- NHMU's Nasutoceratops features in Jurassic World Dominion - 4 minute video interview with curator Randall Irmis
- Battle at Big Rock - 10 minute Jurasic World short film featuring Nasutoceratops
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||

.png.webp)