

The metabotropic glutamate receptors, or mGluRs, are a type of glutamate receptor that are active through an indirect metabotropic process. They are members of the group C family of G-protein-coupled receptors, or GPCRs.[1] Like all glutamate receptors, mGluRs bind with glutamate, an amino acid that functions as an excitatory neurotransmitter.
Function and structure
The mGluRs perform a variety of functions in the central and peripheral nervous systems: For example, they are involved in learning, memory, anxiety, and the perception of pain.[2] They are found in pre- and postsynaptic neurons in synapses of the hippocampus, cerebellum,[3] and the cerebral cortex, as well as other parts of the brain and in peripheral tissues.[4]
Like other metabotropic receptors, mGluRs have seven transmembrane domains that span the cell membrane.[5] Unlike ionotropic receptors, metabotropic glutamate receptors are not ion channels. Instead, they activate biochemical cascades, leading to the modification of other proteins, such as ion channels.[6] This can lead to changes in the synapse's excitability, for example by presynaptic inhibition of neurotransmission,[7] or modulation and even induction of postsynaptic responses.[1][4][5][8]
A dimeric organization of mGluRs is required for signaling induced by agonists.[9]
Classification
Eight different types of mGluRs, labeled mGluR1 to mGluR8 (GRM1 to GRM8), are divided into groups I, II, and III.[1][3][4][8] Receptor types are grouped based on receptor structure and physiological activity.[2] The mGluRs are further divided into subtypes, such as mGluR7a and mGluR7b.
Overview
| Family | Receptors [10][11] | Gene | Mechanism[10] | Function | Agonists & Activators | Antagonists | Synapse site |
|---|---|---|---|---|---|---|---|
| Group I | mGluR1 | GRM1 | Gq, ↑Na+,[4] ↑K+,[4] ↓glutamate[8] |
|
|
mainly postsynaptic[14] | |
| mGluR5 | GRM5 | Gq, ↑Na+,[4] ↑K+,[4] ↓glutamate[8] | |||||
| Group II | mGluR2 | GRM2 | Gi/G0 |
|
mainly presynaptic[14] | ||
| mGluR3 | GRM3 | Gi/G0 | |||||
| Group III | mGluR4 | GRM4 | Gi/G0 |
|
mainly presynaptic[14] | ||
| mGluR6 | GRM6 | Gi/G0 | |||||
| mGluR7 | GRM7 | Gi/G0 | |||||
| mGluR8 | GRM8 | Gi/G0 | |||||
Group I
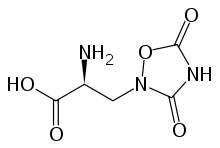
The mGluRs in group I, including mGluR1 and mGluR5, are stimulated most strongly by the excitatory amino acid analog L-quisqualic acid.[4][16] Stimulating the receptors causes the associated enzyme phospholipase C to hydrolyze phosphoinositide phospholipids in the cell's plasma membrane.[1][4][8] This leads to the formation of inositol 1,4,5-trisphosphate (IP3) and diacyl glycerol. Due to its hydrophilic character, IP3 can travel to the endoplasmic reticulum, where it induces, via fixation on its receptor, the opening of calcium channels increasing in this way the cytosolic calcium concentrations. The lipophilic diacylglycerol remains in the membrane, acting as a cofactor for the activation of protein kinase C.
These receptors are also associated with Na+ and K+ channels.[4] Their action can be excitatory, increasing conductance, causing more glutamate to be released from the presynaptic cell, but they also increase inhibitory postsynaptic potentials, or IPSPs.[4] They can also inhibit glutamate release and can modulate voltage-dependent calcium channels.[8]
Group I mGluRs, but not other groups, are activated by 3,5-dihydroxyphenylglycine (DHPG),[14] a fact that is useful to experimenters because it allows them to isolate and identify them.
Group II and Group III
The receptors in group II, including mGluRs 2 and 3, and group III, including mGluRs 4, 6, 7, and 8, (with some exceptions) prevent the formation of cyclic adenosine monophosphate, or cAMP, by activating a G protein that inhibits the enzyme adenylyl cyclase, which forms cAMP from ATP.[1][3][4][17] These receptors are involved in presynaptic inhibition,[8] and do not appear to affect postsynaptic membrane potential by themselves. Receptors in groups II and III reduce the activity of postsynaptic potentials, both excitatory and inhibitory, in the cortex.[4]
The chemicals 2-(2,3-dicarboxycyclopropyl)glycine (DCG-IV) and eglumegad activate only group II mGluRs, while 2-amino-4-phosphonobutyrate (L-AP4) activates only group III mGluRs.[14] Several subtype-selective positive allosteric modulators that activate only the mGlu2 subtype, such as Biphenylindanone A, have also now been developed.
LY-341,495 and MGS-0039 are drugs that act as a selective antagonist blocking both of the group II metabotropic glutamate receptors, mGluR2 and mGluR3.[18] RO4491533 acts as a negative allosteric modulator of mGluR2 and mGluR3.[19]
Localization
Different types of mGluRs are distributed differently in cells. For example, one study found that Group I mGluRs are located mostly on postsynaptic parts of cells, while groups II and III are mostly located on presynaptic elements,[14] though they have been found on both pre- and postsynaptic membranes.[8]
Also, different mGluR subtypes are found predominantly in different parts of the body. For example, mGluR4 is located only in the brain, in locations such as the thalamus, hypothalamus and caudate nucleus.[20] All mGluRs except mGluR6 are thought to exist in the hippocampus and entorhinal cortex.[14]
Roles
It is thought that mGluRs play a role in a variety of different functions.
Modulation of other receptors
Metabotropic glutamate receptors are known to act as modulators of (affect the activity of) other receptors. For example, group I mGluRs are known to increase the activity of N-methyl-D-aspartate receptors (NMDARs),[12][13] a type of ion channel-linked receptor that is central in a neurotoxic process called excitotoxicity. Proteins called PDZ proteins frequently anchor mGluRs near enough to NMDARs to modulate their activity.[21]
It has been suggested that mGluRs may act as regulators of neurons' vulnerability to excitotoxicity (a deadly neurochemical process involving glutamate receptor overactivation) through their modulation of NMDARs, the receptor most involved in that process.[22] Excessive amounts of N-methyl-D-aspartate (NMDA), the selective specific agonist of NMDARs, has been found to cause more damage to neurons in the presence of group I mGluR agonists.[23] On the other hand, agonists of group II[24] and III mGluRs reduce NMDAR activity.[15]
Group II[25] and III[23] mGluRs tend to protect neurons from excitotoxicity,[15][26][27] possibly by reducing the activity of NMDARs.
Metabotropic glutamate receptors are also thought to affect dopaminergic and adrenergic neurotransmission.[28]
Role in plasticity
Like other glutamate receptors, mGluRs have been shown to be involved in synaptic plasticity[1][8] and in neurotoxicity and neuroprotection.[29][30]
They participate in long term potentiation and long term depression, and they are removed from the synaptic membrane in response to agonist binding.[17]
Roles in disease
Since metabotropic glutamate receptors are involved in a variety of functions, abnormalities in their expression can contribute to disease. For example, studies with mutant mice have suggested that mutations in expression of mGluR1 may be involved in the development of certain types of cancer.[31] In addition, manipulating mGluRs can be useful in treating some conditions. For example, clinical trial suggested that an mGlu2/3 agonist, LY354740, was effective in the treatment of generalized anxiety disorder.[32] Also, some researchers have suggested that activation of mGluR4 could be used as a treatment for Parkinson's disease.[33] Most recently, Group I mGluRs, have been implicated in the pathogenesis of Fragile X, a type of autism,[34] and a number of studies are currently testing the therapeutic potential of drugs that modify these receptors.[35] There is also growing evidence that group II metabotropic glutamate receptor agonists may play a role in the treatment of schizophrenia. Schizophrenia is associated with deficits in cortical inhibitory interneurons that release GABA and synaptic abnormalities associated with deficits in NMDA receptor function.[36] These inhibitory deficits may impair cortical function via cortical disinhibition and asynchrony.[37] The drug LY354740 (also known as Eglumegad, an mGlu2/3 agonist) was shown to attenuate physiologic and cognitive abnormalities in animal and human studies of NMDA receptor antagonist and serotonergic hallucinogen effects,[38][39][40][41] supporting the subsequent clinical evidence of efficacy for an mGluR2/3 agonist in the treatment of schizophrenia.[42] The same drug has been shown to interfere in the hypothalamic–pituitary–adrenal axis, with chronic oral administration of this drug leading to markedly reduced baseline cortisol levels in bonnet macaques (Macaca radiata); acute infusion of LY354740 resulted in a marked diminution of yohimbine-induced stress response in those animals.[43] LY354740 has also been demonstrated to act on the metabotropic glutamate receptor 3 (GRM3) of human adrenocortical cells, downregulating aldosterone synthase, CYP11B1, and the production of adrenal steroids (i.e. aldosterone and cortisol).[44]
History
The first demonstration that glutamate could induce the formation of molecules belonging to a major second messenger system was in 1985, when it was shown that it could stimulate the formation of inositol phosphates.[45] This finding allowed in 1987 to yield an explanation for oscillatory ionic glutamate responses and to provide further evidence for the existence of metabotropic glutamate receptors.[46] In 1991 the first metabotropic glutamate receptor of the seven transmembrane domain family was cloned.[47] More recent reports on ionotropic glutamate receptors able to couple to metabotropic transduction systems[48][49] suggest that metabotropic responses of glutamate might not be limited to seven transmembrane domain metabotropic glutamate receptors.
References
- 1 2 3 4 5 6 Bonsi P, Cuomo D, De Persis C, Centonze D, Bernardi G, Calabresi P, Pisani A (2005). "Modulatory action of metabotropic glutamate receptor (mGluR) 5 on mGluR1 function in striatal cholinergic interneurons". Neuropharmacology. 49. 49 (Suppl 1): 104–13. doi:10.1016/j.neuropharm.2005.05.012. PMID 16005029. S2CID 25980146.
- 1 2 Ohashi H, Maruyama T, Higashi-Matsumoto H, Nomoto T, Nishimura S, Takeuchi Y (2002). "A novel binding assay for metabotropic glutamate receptors using [3H] L-quisqualic acid and recombinant receptors" (PDF). Zeitschrift für Naturforschung C. 57 (3–4): 348–55. doi:10.1515/znc-2002-3-425. PMID 12064739. S2CID 23172710. Archived from the original (subscription required) on 2005-10-27. Retrieved 2005-10-08.
- 1 2 3 Hinoi E, Ogita K, Takeuchi Y, Ohashi H, Maruyama T, Yoneda Y (March 2001). "Characterization with [3H]quisqualate of group I metabotropic glutamate receptor subtype in rat central and peripheral excitable tissues". Neurochemistry International. 38 (3): 277–85. doi:10.1016/S0197-0186(00)00075-9. PMID 11099787. S2CID 19402633.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 Chu Z, Hablitz JJ (October 2000). "Quisqualate induces an inward current via mGluR activation in neocortical pyramidal neurons". Brain Research. 879 (1–2): 88–92. doi:10.1016/S0006-8993(00)02752-9. PMID 11011009. S2CID 16433806.
- 1 2 Platt SR (March 2007). "The role of glutamate in central nervous system health and disease--a review". Veterinary Journal. 173 (2): 278–86. doi:10.1016/j.tvjl.2005.11.007. PMID 16376594.
- ↑ Gabriel L, Lvov A, Orthodoxou D, Rittenhouse AR, Kobertz WR, Melikian HE (September 2012). "The acid-sensitive, anesthetic-activated potassium leak channel, KCNK3, is regulated by 14-3-3β-dependent, protein kinase C (PKC)-mediated endocytic trafficking". The Journal of Biological Chemistry. 287 (39): 32354–66. doi:10.1074/jbc.M112.391458. PMC 3463364. PMID 22846993.
- ↑ Sladeczek F., Momiyama A.,Takahashi T. (1992). "Presynaptic inhibitory action of metabotropic glutamate receptor agonist on excitatory transmission in visual cortical neurons". Proc. Roy. Soc. Lond. B 1993 253, 297-303.
- 1 2 3 4 5 6 7 8 9 Endoh T (October 2004). "Characterization of modulatory effects of postsynaptic metabotropic glutamate receptors on calcium currents in rat nucleus tractus solitarius". Brain Research. 1024 (1–2): 212–24. doi:10.1016/j.brainres.2004.07.074. PMID 15451384. S2CID 37860848.
- ↑ El Moustaine D, Granier S, Doumazane E, Scholler P, Rahmeh R, Bron P, Mouillac B, Banères JL, Rondard P, Pin JP (October 2012). "Distinct roles of metabotropic glutamate receptor dimerization in agonist activation and G-protein coupling". Proceedings of the National Academy of Sciences of the United States of America. 109 (40): 16342–7. Bibcode:2012PNAS..10916342E. doi:10.1073/pnas.1205838109. PMC 3479612. PMID 22988116.
- 1 2 If not otherwise specified in table:TABLE 1 Classification of the metabotropic glutamate (mGlu) receptors From the following article:
- ↑ Swanson CJ, Bures M, Johnson MP, Linden AM, Monn JA, Schoepp DD (February 2005). "Metabotropic glutamate receptors as novel targets for anxiety and stress disorders". Nature Reviews. Drug Discovery. 4 (2): 131–44. doi:10.1038/nrd1630. PMID 15665858. S2CID 15037387.
- 1 2 Skeberdis VA, Lan J, Opitz T, Zheng X, Bennett MV, Zukin RS (June 2001). "mGluR1-mediated potentiation of NMDA receptors involves a rise in intracellular calcium and activation of protein kinase C". Neuropharmacology. 40 (7): 856–65. doi:10.1016/S0028-3908(01)00005-3. PMID 11378156. S2CID 19635322.
- 1 2 Lea PM, Custer SJ, Vicini S, Faden AI (September 2002). "Neuronal and glial mGluR5 modulation prevents stretch-induced enhancement of NMDA receptor current". Pharmacology Biochemistry and Behavior. 73 (2): 287–98. doi:10.1016/S0091-3057(02)00825-0. PMID 12117582. S2CID 20086782.
- 1 2 3 4 5 6 7 Shigemoto R, Kinoshita A, Wada E, Nomura S, Ohishi H, Takada M, Flor PJ, Neki A, Abe T, Nakanishi S, Mizuno N (October 1997). "Differential presynaptic localization of metabotropic glutamate receptor subtypes in the rat hippocampus". The Journal of Neuroscience. 17 (19): 7503–22. doi:10.1523/JNEUROSCI.17-19-07503.1997. PMC 6573434. PMID 9295396.
- 1 2 3 4 Ambrosini A, Bresciani L, Fracchia S, Brunello N, Racagni G (May 1995). "Metabotropic glutamate receptors negatively coupled to adenylate cyclase inhibit N-methyl-D-aspartate receptor activity and prevent neurotoxicity in mesencephalic neurons in vitro" (abstract). Molecular Pharmacology. 47 (5): 1057–64. PMID 7746273.
- ↑ Bates B, Xie Y, Taylor N, Johnson J, Wu L, Kwak S, Blatcher M, Gulukota K, Paulsen JE (December 2002). "Characterization of mGluR5R, a novel, metabotropic glutamate receptor 5-related gene". Brain Research. Molecular Brain Research. 109 (1–2): 18–33. doi:10.1016/S0169-328X(02)00458-8. PMID 12531512.
- 1 2 MRC (Medical Research Council), Glutamate receptors: Structures and functions. Archived 2007-09-15 at the Wayback Machine, University of Bristol Centre for Synaptic Plasticity (2003). Retrieved January 20, 2008.
- ↑ Koike H, Iijima M, Chaki S (December 2011). "Involvement of the mammalian target of rapamycin signaling in the antidepressant-like effect of group II metabotropic glutamate receptor antagonists". Neuropharmacology. 61 (8): 1419–23. doi:10.1016/j.neuropharm.2011.08.034. PMID 21903115. S2CID 23923192.
- ↑ Campo B, Kalinichev M, Lambeng N, El Yacoubi M, Royer-Urios I, Schneider M, Legrand C, Parron D, Girard F, Bessif A, Poli S, Vaugeois JM, Le Poul E, Celanire S (December 2011). "Characterization of an mGluR2/3 negative allosteric modulator in rodent models of depression". Journal of Neurogenetics. 25 (4): 152–66. doi:10.3109/01677063.2011.627485. PMID 22091727. S2CID 207440972.
- ↑ InterPro. InterPro: IPR001786 Metabotropic glutamate receptor 4. Retrieved on January 20, 2008.
- ↑ Tu JC, Xiao B, Naisbitt S, Yuan JP, Petralia RS, Brakeman P, Doan A, Aakalu VK, Lanahan AA, Sheng M, Worley PF (July 1999). "Coupling of mGluR/Homer and PSD-95 complexes by the Shank family of postsynaptic density proteins". Neuron. 23 (3): 583–92. doi:10.1016/S0896-6273(00)80810-7. PMID 10433269. S2CID 16429070.
- ↑ Baskys A, Blaabjerg M (March 2005). "Understanding regulation of nerve cell death by mGluRs as a method for development of successful neuroprotective strategies". Journal of the Neurological Sciences. 229–230: 201–9. doi:10.1016/j.jns.2004.11.028. PMID 15760640. S2CID 39812016.
- 1 2 Bruno V, Copani A, Knöpfel T, Kuhn R, Casabona G, Dell'Albani P, Condorelli DF, Nicoletti F (August 1995). "Activation of metabotropic glutamate receptors coupled to inositol phospholipid hydrolysis amplifies NMDA-induced neuronal degeneration in cultured cortical cells". Neuropharmacology. 34 (8): 1089–98. doi:10.1016/0028-3908(95)00077-J. PMID 8532158. S2CID 23848439.
- ↑ Buisson A, Yu SP, Choi DW (January 1996). "DCG-IV selectively attenuates rapidly triggered NMDA-induced neurotoxicity in cortical neurons". The European Journal of Neuroscience. 8 (1): 138–43. doi:10.1111/j.1460-9568.1996.tb01174.x. PMID 8713457. S2CID 24122616.
- ↑ Bruno V, Battaglia G, Copani A, Giffard RG, Raciti G, Raffaele R, Shinozaki H, Nicoletti F (September 1995). "Activation of class II or III metabotropic glutamate receptors protects cultured cortical neurons against excitotoxic degeneration". The European Journal of Neuroscience. 7 (9): 1906–13. doi:10.1111/j.1460-9568.1995.tb00712.x. PMID 8528465. S2CID 10181885.
- ↑ Allen JW, Ivanova SA, Fan L, Espey MG, Basile AS, Faden AI (July 1999). "Group II metabotropic glutamate receptor activation attenuates traumatic neuronal injury and improves neurological recovery after traumatic brain injury" (abstract). The Journal of Pharmacology and Experimental Therapeutics. 290 (1): 112–20. PMID 10381766.
- ↑ Faden AI, Ivanova SA, Yakovlev AG, Mukhin AG (December 1997). "Neuroprotective effects of group III mGluR in traumatic neuronal injury". Journal of Neurotrauma. 14 (12): 885–95. doi:10.1089/neu.1997.14.885. PMID 9475370.
- ↑ Wang J-Q, Brownell A-L (2007). "Development of metabotropic glutamate receptor ligands for neuroimaging". Current Medical Imaging Reviews. 3 (3): 186–205. doi:10.2174/157340507781387059.
- ↑ Siliprandi R, Lipartiti M, Fadda E, Sautter J, Manev H (August 1992). "Activation of the glutamate metabotropic receptor protects retina against N-methyl-D-aspartate toxicity". European Journal of Pharmacology. 219 (1): 173–4. doi:10.1016/0014-2999(92)90598-X. PMID 1397046.
- ↑ Baskys A, Fang L, Bayazitov I (August 2005). "Activation of neuroprotective pathways by metabotropic group I glutamate receptors: a potential target for drug discovery?". Annals of the New York Academy of Sciences. 1053 (1): 55–73. Bibcode:2005NYASA1053...55B. doi:10.1196/annals.1344.006. PMID 16179509.
- ↑ Namkoong J, Shin SS, Lee HJ, Marín YE, Wall BA, Goydos JS, Chen S (March 2007). "Metabotropic glutamate receptor 1 and glutamate signaling in human melanoma". Cancer Research. 67 (5): 2298–305. doi:10.1158/0008-5472.CAN-06-3665. PMID 17332361.
- ↑ Dunayevich E, Erickson J, Levine L, Landbloom R, Schoepp DD, Tollefson GD (June 2008). "Efficacy and tolerability of an mGlu2/3 agonist in the treatment of generalized anxiety disorder". Neuropsychopharmacology. 33 (7): 1603–10. doi:10.1038/sj.npp.1301531. PMID 17712352.
- ↑ Marino MJ, Williams DL, O'Brien JA, Valenti O, McDonald TP, Clements MK, Wang R, DiLella AG, Hess JF, Kinney GG, Conn PJ (November 2003). "Allosteric modulation of group III metabotropic glutamate receptor 4: a potential approach to Parkinson's disease treatment". Proceedings of the National Academy of Sciences of the United States of America. 100 (23): 13668–73. Bibcode:2003PNAS..10013668M. doi:10.1073/pnas.1835724100. PMC 263871. PMID 14593202.
- ↑ Dölen G, Osterweil E, Rao BS, Smith GB, Auerbach BD, Chattarji S, Bear MF (December 2007). "Correction of fragile X syndrome in mice". Neuron. 56 (6): 955–62. doi:10.1016/j.neuron.2007.12.001. PMC 2199268. PMID 18093519.
- ↑ Dölen G, Carpenter RL, Ocain TD, Bear MF (July 2010). "Mechanism-based approaches to treating fragile X". Pharmacology & Therapeutics. 127 (1): 78–93. doi:10.1016/j.pharmthera.2010.02.008. PMID 20303363.
- ↑ Krystal JH, D'Souza DC, Mathalon D, Perry E, Belger A, Hoffman R (September 2003). "NMDA receptor antagonist effects, cortical glutamatergic function, and schizophrenia: toward a paradigm shift in medication development". Psychopharmacology. 169 (3–4): 215–33. doi:10.1007/s00213-003-1582-z. PMID 12955285. S2CID 25800929.
- ↑ Ford JM, Krystal JH, Mathalon DH (July 2007). "Neural synchrony in schizophrenia: from networks to new treatments". Schizophrenia Bulletin. 33 (4): 848–52. doi:10.1093/schbul/sbm062. PMC 2632315. PMID 17567628.
- ↑ Homayoun H, Jackson ME, Moghaddam B (April 2005). "Activation of metabotropic glutamate 2/3 receptors reverses the effects of NMDA receptor hypofunction on prefrontal cortex unit activity in awake rats". Journal of Neurophysiology. 93 (4): 1989–2001. doi:10.1152/jn.00875.2004. PMID 15590730. S2CID 5472650.
- ↑ Moghaddam B, Adams BW (August 1998). "Reversal of phencyclidine effects by a group II metabotropic glutamate receptor agonist in rats". Science. 281 (5381): 1349–52. Bibcode:1998Sci...281.1349M. doi:10.1126/science.281.5381.1349. PMID 9721099.
- ↑ Krystal JH, Abi-Saab W, Perry E, D'Souza DC, Liu N, Gueorguieva R, McDougall L, Hunsberger T, Belger A, Levine L, Breier A (April 2005). "Preliminary evidence of attenuation of the disruptive effects of the NMDA glutamate receptor antagonist, ketamine, on working memory by pretreatment with the group II metabotropic glutamate receptor agonist, LY354740, in healthy human subjects". Psychopharmacology. 179 (1): 303–9. doi:10.1007/s00213-004-1982-8. PMID 15309376. S2CID 29731346.
- ↑ Aghajanian GK, Marek GJ (March 2000). "Serotonin model of schizophrenia: emerging role of glutamate mechanisms". Brain Research. Brain Research Reviews. 31 (2–3): 302–12. doi:10.1016/S0165-0173(99)00046-6. PMID 10719157. S2CID 13040014.
- ↑ Patil ST, Zhang L, Martenyi F, Lowe SL, Jackson KA, Andreev BV, Avedisova AS, Bardenstein LM, Gurovich IY, Morozova MA, Mosolov SN, Neznanov NG, Reznik AM, Smulevich AB, Tochilov VA, Johnson BG, Monn JA, Schoepp DD (September 2007). "Activation of mGlu2/3 receptors as a new approach to treat schizophrenia: a randomized Phase 2 clinical trial". Nature Medicine. 13 (9): 1102–7. doi:10.1038/nm1632. PMID 17767166. S2CID 6417333.
- ↑ Coplan JD, Mathew SJ, Smith EL, Trost RC, Scharf BA, Martinez J, Gorman JM, Monn JA, Schoepp DD, Rosenblum LA (July 2001). "Effects of LY354740, a novel glutamatergic metabotropic agonist, on nonhuman primate hypothalamic-pituitary-adrenal axis and noradrenergic function". CNS Spectrums. 6 (7): 607–12, 617. doi:10.1017/S1092852900002157. PMID 15573025. S2CID 6029856.
- ↑ Felizola SJ, Nakamura Y, Satoh F, Morimoto R, Kikuchi K, Nakamura T, Hozawa A, Wang L, Onodera Y, Ise K, McNamara KM, Midorikawa S, Suzuki S, Sasano H (January 2014). "Glutamate receptors and the regulation of steroidogenesis in the human adrenal gland: the metabotropic pathway". Molecular and Cellular Endocrinology. 382 (1): 170–7. doi:10.1016/j.mce.2013.09.025. PMID 24080311. S2CID 3357749.
- ↑ Sladeczek F, Pin JP, Récasens M, Bockaert J, Weiss S (1985). "Glutamate stimulates inositol phosphate formation in striatal neurones". Nature. 317 (6039): 717–9. Bibcode:1985Natur.317..717S. doi:10.1038/317717a0. PMID 2865680. S2CID 3192974.
- ↑ Sugiyama H, Ito I, Hirono C (1987). "A new type of glutamate receptor linked to inositol phospholipid metabolism". Nature. 325 (6104): 531–3. Bibcode:1987Natur.325..531S. doi:10.1038/325531a0. PMID 2880300. S2CID 4364831.
- ↑ Masu M, Tanabe Y, Tsuchida K, Shigemoto R, Nakanishi S (February 1991). "Sequence and expression of a metabotropic glutamate receptor". Nature. 349 (6312): 760–5. Bibcode:1991Natur.349..760M. doi:10.1038/349760a0. PMID 1847995. S2CID 4325501.
- ↑ Dingledine R, Borges K, Bowie D, Traynelis SF (March 1999). "The glutamate receptor ion channels". Pharmacological Reviews. 51 (1): 7–61. PMID 10049997.
- ↑ Wang Y, Small DL, Stanimirovic DB, Morley P, Durkin JP (October 1997). "AMPA receptor-mediated regulation of a Gi-protein in cortical neurons". Nature. 389 (6650): 502–4. Bibcode:1997Natur.389..502W. doi:10.1038/39062. PMID 9333240. S2CID 205026915.
Further reading
- Jin, Chaobin; Ma, Shutao (2017). "Recent advances in the medicinal chemistry of group II and group III mGlu receptors". Med. Chem. Commun. 8 (3): 501–515. doi:10.1039/C6MD00612D. ISSN 2040-2503. PMC 6072351. PMID 30108768.
- O'Brien DE, Conn PJ (2016). "Neurobiological Insights from mGlu Receptor Allosteric Modulation". Int. J. Neuropsychopharmacol. 19 (5): pyv133. doi:10.1093/ijnp/pyv133. PMC 4886670. PMID 26647381.
- Gregory KJ, Conn PJ (2015). "Molecular Insights into Metabotropic Glutamate Receptor Allosteric Modulation". Mol. Pharmacol. 88 (1): 188–202. doi:10.1124/mol.114.097220. PMC 4468636. PMID 25808929.
External links
- Metabotropic+Glutamate+Receptors at the U.S. National Library of Medicine Medical Subject Headings (MeSH)