
Jasmonate (JA) and its derivatives are lipid-based plant hormones that regulate a wide range of processes in plants, ranging from growth and photosynthesis to reproductive development. In particular, JAs are critical for plant defense against herbivory and plant responses to poor environmental conditions and other kinds of abiotic and biotic challenges.[1] Some JAs can also be released as volatile organic compounds (VOCs) to permit communication between plants in anticipation of mutual dangers.[2]
History
The isolation of methyl jasmonate (MeJa) from jasmine oil derived from Jasminum grandiflorum led to the discovery of the molecular structure of jasmonates and their name in 1962[3][4] while jasmonic acid itself was isolated from Lasiodiplodia theobromae by Alderidge et al in 1971.[4]
Biosynthesis


Biosynthesis is reviewed by Acosta and Farmer 2010, Wasternack and Hause 2013, and Wasternack and Song 2017.[4] Jasmonates (JA) are oxylipins, i.e. derivatives of oxygenated fatty acid. They are biosynthesized from linolenic acid in chloroplast membranes. Synthesis is initiated with the conversion of linolenic acid to 12-oxo-phytodienoic acid (OPDA), which then undergoes a reduction and three rounds of oxidation to form (+)-7-iso-JA, jasmonic acid. Only the conversion of linolenic acid to OPDA occurs in the chloroplast; all subsequent reactions occur in the peroxisome.[5]
JA itself can be further metabolized into active or inactive derivatives. Methyl JA (MeJA) is a volatile compound that is potentially responsible for interplant communication. JA conjugated with amino acid isoleucine (Ile) results in JA-Ile ((+)-7-iso-jasmonoyl-L-isoleucine), which Fonseca et al 2009 finds is involved in most JA signaling[6] - see also the review by Katsir et al 2008.[5] However Van Poecke & Dicke 2003 finds Arabidopsis's emission of volatiles to not require JA-Ile, nor VanDoorn et al 2011 for Solanum nigrum's herbivore resistance.[6] JA undergoes decarboxylation to give cis-jasmone.
Function
Although jasmonate (JA) regulates many different processes in the plant, its role in wound response is best understood. Following mechanical wounding or herbivory, JA biosynthesis is rapidly activated, leading to expression of the appropriate response genes. For example, in the tomato, wounding produces defense molecules that inhibit leaf digestion in guts of insects. Another indirect result of JA signaling is the volatile emission of JA-derived compounds. MeJA on leaves can travel airborne to nearby plants and elevate levels of transcripts related to wound response.[1] In general, this emission can further upregulate JA biosynthesis and cell signaling, thereby inducing nearby plants to prime their defenses in case of herbivory.
JAs have also been implicated in cell death and leaf senescence. JA can interact with many kinases and transcription factors associated with senescence. JA can also induce mitochondrial death by inducing the accumulation of reactive oxygen species (ROSs). These compounds disrupt mitochondria membranes and compromise the cell by causing apoptosis, or programmed cell death. JAs' roles in these processes are suggestive of methods by which the plant defends itself against biotic challenges and limits the spread of infections.[7]
JA and its derivatives have also been implicated in plant development, symbiosis, and a host of other processes included in the list below.
- By studying mutants overexpressing JA, one of the earliest discoveries made was that JA inhibits root growth. The mechanism behind this event is still not understood, but mutants in the COI1-dependent signaling pathway tend to show reduced inhibition, demonstrating that the COI1 pathway is somehow necessary for inhibiting root growth.[8][9]
- JA plays many roles in flower development. Mutants in JA synthesis or in JA signaling in Arabidopsis present with male sterility, typically due to delayed development. The same genes promoting male fertility in Arabidopsis promote female fertility in tomatoes. Overexpression of 12-OH-JA can also delay flowering.[9]
- JA and MeJA inhibit the germination of nondormant seeds and stimulate the germination of dormant seeds.[10]
- High levels of JA encourage the accumulation of storage proteins; genes encoding vegetative storage proteins are JA responsive. Specifically, tuberonic acid, a JA derivative, induces the formation of tubers.[11][12]
- JAs also play a role in symbiosis between plants and microorganisms; however, its precise role is still unclear. JA currently appears to regulate signal exchange and nodulation regulation between legumes and rhizobium. On the other hand, elevated JA levels appear to regulate carbohydrate partitioning and stress tolerance in mycorrhizal plants.[13]

- JAs have been implicated in the development of carnivorous plants such as the Venus flytrap. Research suggests that evolutionary repurposing of the jasmonate signaling pathway, which mediates defense against herbivores in noncarnivorous plants, has supported the evolution of plant carnivory. Jasmonates can be used to signal the closing of traps and to control the release of enzymes and nutrient transporters which are used in plant digestion. However, not all carnivorous plants rely on the jasmonate pathway in the same way. Butterworts differ significantly from Venus flytraps and sundews, and may have developed methods of regulating digestive enzymes that are JA-independent.[14][15]
Role in pathogenesis
Pseudomonas syringae causes bacterial speck disease in tomatoes by hijacking the plant's jasmonate (JA) signaling pathway. This bacteria utilizes a type III secretion system to inject a cocktail of viral effector proteins into host cells.
One of the molecules included in this mixture is the phytotoxin coronatine (COR). JA-insensitive plants are highly resistant to P. syringae and unresponsive to COR; additionally, applying MeJA was sufficient to rescue virulence in COR mutant bacteria. Infected plants also expressed downstream JA and wound response genes but repressed levels of pathogenesis-related (PR) genes. All these data suggest COR acts through the JA pathway to invade host plants. Activation of a wound response is hypothesized to come at the expense of pathogen defense. By activating the JA wound response pathway, P. syringae could divert resources from its host's immune system and infect more effectively.[16]
Plants produce N-acylamides that confer resistance to necrotrophic pathogens by activating JA biosynthesis and signalling. Arachidonic acid (AA), the counterpart of the JA precursor α-LeA occurring in metazoan species but not in plants, is perceived by plants and acts through an increase in JA levels concomitantly with resistance to necrotrophic pathogens. AA is an evolutionarily conserved signalling molecule that acts in plants in response to stress similar to that in animal systems.[17]
Cross talk with other defense pathways
While the jasmonate (JA) pathway is critical for wound response, it is not the only signaling pathway mediating defense in plants. To build an optimal yet efficient defense, the different defense pathways must be capable of cross talk to fine-tune and specify responses to abiotic and biotic challenges.
One of the best studied examples of JA cross talk occurs with salicylic acid (SA). SA, a hormone, mediates defense against pathogens by inducing both the expression of pathogenesis-related genes and systemic acquired resistance (SAR), in which the whole plant gains resistance to a pathogen after localized exposure to it.
Wound and pathogen response appear to be interact negatively. For example, silencing phenylalanine ammonia lyase (PAL), an enzyme synthesizing precursors to SA, reduces SAR but enhances herbivory resistance against insects. Similarly, overexpression of PAL enhances SAR but reduces wound response after insect herbivory.[18] Generally, it has been found that pathogens living in live plant cells are more sensitive to SA-induced defenses, while herbivorous insects and pathogens that derive benefit from cell death are more susceptible to JA defenses. Thus, this trade-off in pathways optimizes defense and saves plant resources.[19]
Cross talk also occurs between JA and other plant hormone pathways, such as those of abscisic acid (ABA) and ethylene (ET). These interactions similarly optimize defense against pathogens and herbivores of different lifestyles. For example, MYC2 activity can be stimulated by both JA and ABA pathways, allowing it to integrate signals from both pathways. Other transcription factors such as ERF1 arise as a result of JA and ET signaling. All these molecules can act in combination to activate specific wound response genes.[19]
Finally, cross talk is not restricted for defense: JA and ET interactions are critical in development as well, and a balance between the two compounds is necessary for proper apical hook development in Arabidopsis seedlings. Still, further research is needed to elucidate the molecules regulating such cross talk.[18]
Mechanism of signaling
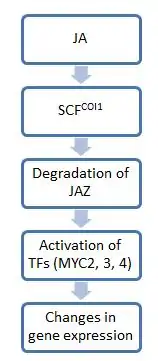
In general, the steps in jasmonate (JA) signaling mirror that of auxin signaling: the first step comprises E3 ubiquitin ligase complexes, which tag substrates with ubiquitin to mark them for degradation by proteasomes. The second step utilizes transcription factors to effect physiological changes. One of the key molecules in this pathway is JAZ, which serves as the on-off switch for JA signaling. In the absence of JA, JAZ proteins bind to downstream transcription factors and limit their activity. However, in the presence of JA or its bioactive derivatives, JAZ proteins are degraded, freeing transcription factors for expression of genes needed in stress responses.[20]
Because JAZ did not disappear in null coi1 mutant plant backgrounds, protein COI1 was shown to mediate JAZ degradation. COI1 belongs to the family of highly conserved F-box proteins, and it recruits substrates for the E3 ubiquitin ligase SCFCOI1. The complexes that ultimately form are known as SCF complexes.[21] These complexes bind JAZ and target it for proteasomal degradation. However, given the large spectrum of JA molecules, not all JA derivatives activate this pathway for signaling, and the range of those participating in this pathway is unknown.[5] Thus far, only JA-Ile has been shown to be necessary for COI1-mediated degradation of JAZ11. JA-Ile and structurally related derivatives can bind to COI1-JAZ complexes and promote ubiquitination and thus degradation of the latter.[5]
This mechanistic model raises the possibility that COI1 serves as an intracellular receptor for JA signals. Recent research has confirmed this hypothesis by demonstrating that the COI1-JAZ complex acts as a co-receptor for JA perception. Specifically, JA-Ile binds both to a ligand-binding pocket in COI1 and to a 20 amino-acid stretch of the conserved Jas motif in JAZ. This JAZ residue acts as a plug for the pocket in COI1, keeping JA-Ile bound in the pocket. Additionally, Sheard et al 2010[22] co-purified and subsequently removed inositol pentakisphosphate (InsP5) from COI1, demonstrating InsP5 to be a necessary component of the co-receptor and playing a role in potentiating the co-receptor complex. Sheard's results may show varying binding specificity for various SCFCOI1-InsP5-JAZ complexes.[6]
Once freed from JAZ, transcription factors can activate genes needed for a specific JA response. The best-studied transcription factors acting in this pathway belong to the MYC family of transcription factors, which are characterized by a basic helix-loop-helix (bHLH) DNA binding motif. These factors (of which there are three, MYC2, 3, and 4) tend to act additively. For example, a plant that has only lost one myc becomes more susceptible to insect herbivory than a normal plant. A plant that has lost all three will be as susceptible to damage as coi1 mutants, which are completely unresponsive to JA and cannot mount a defense against herbivory. However, while all these MYC molecules share functions, they vary greatly in expression patterns and transcription functions. For instance, MYC2 has a greater effect on root growth compared to MYC3 or MYC4.[8]
Additionally, MYC2 will loop back and regulate JAZ expression levels, leading to a negative feedback loop.[8] These transcription factors all have different impacts on JAZ levels after JA signaling. JAZ levels in turn affect transcription factor and gene expression levels. In other words, on top of activating different response genes, the transcription factors can vary JAZ levels to achieve specificity in response to JA signals.
References
- 1 2 Farmer, E. E.; Ryan, C. A. (1990). "Interplant communication: airborne methyl jasmonate induces synthesis of proteinase inhibitors in plant leaves". Proc Natl Acad Sci U S A. 87 (19): 7713–7716. Bibcode:1990PNAS...87.7713F. doi:10.1073/pnas.87.19.7713. PMC 54818. PMID 11607107.
- ↑ Baldwin, I. T.; Halitschke, R.; Paschold, A.; von Dahl, C. C.; Preston, C. A. (2006). "Volatile signaling in plant-plant interactions: "talking trees" in the genomics era". Science. 311 (5762): 812–815. Bibcode:2006Sci...311..812B. doi:10.1126/science.1118446. PMID 16469918. S2CID 9260593.
- ↑ Demole E; Lederer, E.; Mercier, D. (1962). "Isolement et détermination de la structure du jasmonate de méthyle, constituant odorant caractéristique de l'essence de jasmin". Helv Chim Acta. 45 (2): 675–85. doi:10.1002/hlca.19620450233.
- 1 2 3 Per, Tasir S.; Khan, M. Iqbal R.; Anjum, Naser A.; Masood, Asim; Hussain, Sofi Javed; Khan, Nafees A. (2018). "Jasmonates in plants under abiotic stresses: Crosstalk with other phytohormones matters". Environmental and Experimental Botany. Elsevier. 145: 104–120. doi:10.1016/j.envexpbot.2017.11.004. ISSN 0098-8472.
- 1 2 3 4 Katsir, L.; Chung, H. S.; Koo, A. J. K.; Howe, G. A. (2008). "Jasmonate signaling: a conserved mechanism of hormone sensing". Current Opinion in Plant Biology. Elsevier. 11 (4): 428–435. doi:10.1016/j.pbi.2008.05.004. PMC 2560989. PMID 18583180.
- 1 2 3 Schuman, Meredith C.; Baldwin, Ian T. (2016-03-11). "The Layers of Plant Responses to Insect Herbivores". Annual Review of Entomology. Annual Reviews. 61 (1): 373–394. doi:10.1146/annurev-ento-010715-023851. ISSN 0066-4170. PMID 26651543. S2CID 24720368.
- ↑ Reinbothe, C; Springer, A; Samol, I; Reinbothe, S (September 2009). "Plant oxylipins: role of jasmonic acid during programmed cell death, defence and leaf senescence". The FEBS Journal. 276 (17): 4666–81. doi:10.1111/j.1742-4658.2009.07193.x. PMID 19663906. S2CID 1349010.
- 1 2 3 Fernandez-Calvo, P.; Chini, A.; Fernandez-Barbero, G.; Chico, J.-M.; Gimenez-Ibanez, S.; Geerinck, J.; Eeckhout, D.; Schweizer, F.; Godoy, M.; Franco-Zorrilla, J. M.; Pauwels, L.; Witters, E.; Puga, M. I.; Paz-Ares, J.; Goossens, A.; Reymond, P.; De Jaeger, G.; Solano, R. (18 February 2011). "The Arabidopsis bHLH Transcription Factors MYC3 and MYC4 Are Targets of JAZ Repressors and Act Additively with MYC2 in the Activation of Jasmonate Responses". The Plant Cell Online. 23 (2): 701–715. doi:10.1105/tpc.110.080788. PMC 3077776. PMID 21335373.
- 1 2 Wasternack, C. (18 May 2007). "Jasmonates: An Update on Biosynthesis, Signal Transduction and Action in Plant Stress Response, Growth and Development". Annals of Botany. 100 (4): 681–697. doi:10.1093/aob/mcm079. PMC 2749622. PMID 17513307.
- ↑ Creelman, Robert A.; Mullet, John E. (1 June 1997). "Biosynthesis and Action of Jasmonates in Plants". Annual Review of Plant Physiology and Plant Molecular Biology. 48 (1): 355–381. doi:10.1146/annurev.arplant.48.1.355. PMID 15012267.
- ↑ Anderson, JM. (1988). "Jasmonic acid-dependent increases in the level of specific polypeptides in soybean suspension cultures and seedlings". Journal of Plant Growth and Regulation. 7 (4): 203–11. doi:10.1007/BF02025263. S2CID 37785073.
- ↑ Pelacho, AM; Mingo-Castel, AM. (1991). "Jasmonic acid induces tuberization of potato stolons cultured in vitro". Plant Physiology. 97 (3): 1253–55. doi:10.1104/pp.97.3.1253. PMC 1081150. PMID 16668517.
- ↑ Hause, Bettina; Schaarschmidt, Sara (1 September 2009). "The role of jasmonates in mutualistic symbioses between plants and soil-born microorganisms". Phytochemistry. 70 (13–14): 1589–1599. doi:10.1016/j.phytochem.2009.07.003. PMID 19700177.
- 1 2 Pain, Stephanie (2 March 2022). "How plants turned predator". Knowable Magazine. doi:10.1146/knowable-030122-1. Retrieved 11 March 2022.
- ↑ Hedrich, Rainer; Fukushima, Kenji (17 June 2021). "On the Origin of Carnivory: Molecular Physiology and Evolution of Plants on an Animal Diet". Annual Review of Plant Biology. 72 (1): 133–153. doi:10.1146/annurev-arplant-080620-010429. ISSN 1543-5008. PMID 33434053. S2CID 231595236. Retrieved 11 March 2022.
- ↑ Zhao, Y; Thilmony, R; Bender, CL; Schaller, A; He, SY; Howe, GA (November 2003). "Virulence systems of Pseudomonas syringae pv. tomato promote bacterial speck disease in tomato by targeting the jasmonate signaling pathway". The Plant Journal. 36 (4): 485–99. doi:10.1046/j.1365-313x.2003.01895.x. PMID 14617079.
- ↑ Wasternack, C.; Hause, B. (June 2013). "Jasmonates: biosynthesis, perception, signal transduction and action in plant stress response, growth and development. An update to the 2007 review in Annals of Botany". Annals of Botany. 111 (6): 1021–1058. doi:10.1093/aob/mct067. ISSN 0305-7364. PMC 3662512. PMID 23558912.
- 1 2 Turner, J. G.; Ellis, C.; Devoto, A. (2002). "The jasmonate signal pathway". Plant Cell. 14 Suppl (7): 153–164. doi:10.1105/tpc.000679. PMC 151253. PMID 12045275.
- 1 2 Koornneef, A.; Pieterse, C. M.J. (1 March 2008). "Cross Talk in Defense Signaling". Plant Physiology. 146 (3): 839–844. doi:10.1104/pp.107.112029. PMC 2259093. PMID 18316638.
- ↑ Chini, A.; Fonseca, S.; Fernandez, G.; Adie, B.; Chico, J. M.; Lorenzo, O.; Garcia-Casado, G.; Lopez-Vidriero, I.; Lozano, F. M.; Ponce, M. R.; Micol, J. L; Solano, R. (2007). "The JAZ family of repressors is the missing link in jasmonate signaling". Nature. 448 (7154): 666–671. Bibcode:2007Natur.448..666C. doi:10.1038/nature06006. PMID 17637675. S2CID 4383741.
- ↑ Devoto, A; Nieto-Rostro, M; Xie, D; Ellis, C; Harmston, R; Patrick, E; Davis, J; Sherratt, L; Coleman, M; Turner, JG (November 2002). "COI1 links jasmonate signalling and fertility to the SCF ubiquitin-ligase complex in Arabidopsis". The Plant Journal. 32 (4): 457–66. doi:10.1046/j.1365-313x.2002.01432.x. PMID 12445118.
- ↑ Sheard, Laura B.; Tan, Xu; Mao, Haibin; Withers, John; Ben-Nissan, Gili; Hinds, Thomas R.; Kobayashi, Yuichi; Hsu, Fong-Fu; Sharon, Michal; Browse, John; He, Sheng Yang; Rizo, Josep; Howe, Gregg A.; Zheng, Ning (6 October 2010). "Jasmonate perception by inositol-phosphate-potentiated COI1–JAZ co-receptor". Nature. 468 (7322): 400–405. Bibcode:2010Natur.468..400S. doi:10.1038/nature09430. PMC 2988090. PMID 20927106.