| Choline acetyltransferase | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||
| EC no. | 2.3.1.6 | ||||||||
| CAS no. | 9012-78-6 | ||||||||
| Databases | |||||||||
| IntEnz | IntEnz view | ||||||||
| BRENDA | BRENDA entry | ||||||||
| ExPASy | NiceZyme view | ||||||||
| KEGG | KEGG entry | ||||||||
| MetaCyc | metabolic pathway | ||||||||
| PRIAM | profile | ||||||||
| PDB structures | RCSB PDB PDBe PDBsum | ||||||||
| Gene Ontology | AmiGO / QuickGO | ||||||||
| |||||||||
| CHAT | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Identifiers | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Aliases | CHAT, CHOACTASE, CMS1A, CMS1A2, CMS6, choline O-acetyltransferase | ||||||||||||||||||||||||||||||||||||||||||||||||||
| External IDs | OMIM: 118490 MGI: 88392 HomoloGene: 40693 GeneCards: CHAT | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Wikidata | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




Choline acetyltransferase (commonly abbreviated as ChAT, but sometimes CAT) is a transferase enzyme responsible for the synthesis of the neurotransmitter acetylcholine. ChAT catalyzes the transfer of an acetyl group from the coenzyme acetyl-CoA to choline, yielding acetylcholine (ACh). ChAT is found in high concentration in cholinergic neurons, both in the central nervous system (CNS) and peripheral nervous system (PNS). As with most nerve terminal proteins, ChAT is produced in the body of the neuron and is transported to the nerve terminal, where its concentration is highest. Presence of ChAT in a nerve cell classifies this cell as a "cholinergic" neuron. In humans, the choline acetyltransferase enzyme is encoded by the CHAT gene.[5]
History
Choline acetyltransferase was first described by David Nachmansohn and A. L. Machado in 1943.[6] A German biochemist, Nachmansohn had been studying the process of nerve impulse conduction and utilization of energy-yielding chemical reactions in cells, expanding upon the works of Nobel laureates Otto Warburg and Otto Meyerhof on fermentation, glycolysis, and muscle contraction. Based on prior research showing that "acetylcholine's actions on structural proteins" were responsible for nerve impulses, Nachmansohn and Machado investigated the origin of acetylcholine.[7]
An enzyme has been extracted from brain and nervous tissue which forms acetylcholine. The formation occurs only in presence of adenosinetriphosphate (ATP). The enzyme is called choline acetylase.
— Nachmanson & Machado, 1943[6]
The acetyl transferase mode of action was unknown at the time of this discovery, however Nachmansohn hypothesized the possibility of acetylphosphate or phosphorylcholine exchanging the phosphate (from ATP) for choline or acetate ion.[6] It was not until 1945 that Coenzyme A (CoA) was discovered simultaneously and independently by three laboratories,[8][9][10] Nachmansohn's being one of these. Subsequently, acetyl-CoA, at the time called “active acetate,” was discovered in 1951.[11] The 3D structure of rat-derived ChAT was not solved until nearly 60 years later, in 2004.[12]
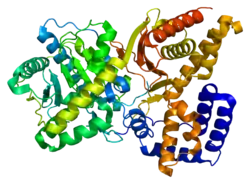
Structure
The 3D structure of ChAT has been solved by X-ray crystallography PDB: 2FY2. Choline is bound in the active site of ChAT by non-covalent interactions between the positively charged amine of choline and the hydroxyl group of Tyr552, in addition to a hydrogen bond between choline's hydroxyl group and a histidine residue, His324.
The choline substrate fits into a pocket in the interior of ChAT, while acetyl-CoA fits into a pocket on the surface of the protein. The 3D crystal structure shows the acetyl group of acetyl-CoA abuts the choline binding pocket – minimizing the distance between acetyl-group donor and receiver.
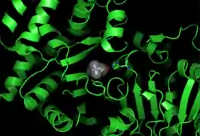
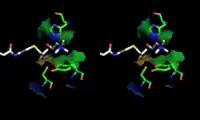
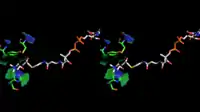
Homology
ChAT is very conserved across the animal genome. Among mammals, in particular, there is very high sequence similarity. Human and cat (Felis catus) ChAT, for example, have 89% sequence identity. Sequence identity with Drosophila is about 30%.[13]
Forms of ChAT
There are two forms of ChAT: Soluble form and membrane-bound form.[14] The soluble form accounts for 80-90% of the total enzyme activity while the membrane-bound form is responsible for the rest of 10-20% activity.[15] However, there has long been a debate on how the latter form of ChAT is bound to the membrane.[16] The membrane-bound form of ChAT is associated with synaptic vesicles.[17]
Common and peripheral isoforms of ChAT
There exist two isoforms of ChAT, both encoded by the same sequence. The common type ChAT (cChAT) is present in both the CNS and PNS. Peripheral type ChAT (pChAT) is preferentially expressed in the PNS in humans, and arises from exon skipping (exons 6–9) during post-transcriptional modification. Therefore, the amino acid sequence is very similar; however, pChAT is missing parts of the sequence present in cChAT. The pChAT isoform was discovered in 2000 based on observations that brain-derived ChAT antibodies failed to stain peripheral cholinergic neurons as they do for those found in the brain. This gene splicing mechanism which leads to cChAT and pChAT differences has been observed in various species, including both vertebrate mammals and invertebrate mollusks, suggesting this mechanism leads to some yet-unidentified evolutionary advantage.
Function
Cholinergic systems are implicated in numerous neurologic functions. Alteration in some cholinergic neurons may account for the disturbances of Alzheimer disease. The protein encoded by this gene synthesizes the neurotransmitter acetylcholine. Acetylcholine acts at two classes of receptors in the central nervous system – muscarinic and nicotinic – which are each implicated in different physiological responses. The role of acetylcholine at the nicotinic receptor is still under investigation. It is likely implicated in the reward/reinforcement pathways, as indicated by the addictive nature of nicotine, which also binds to the nicotinic receptor. The muscarinic action of acetylcholine in the CNS is implicated in learning and memory. The loss of cholinergic innervation in the neocortex has been associated with memory loss, as is evidenced in advanced cases of Alzheimer's disease. In the peripheral nervous system, cholinergic neurons are implicated in the control of visceral functions such as, but not limited to, cardiac muscle contraction and gastrointestinal tract function.

It is often used as an immunohistochemical marker for motor neurons (motoneurons).
Mutations
Mutants of ChAT have been isolated in several species, including C. elegans, Drosophila, and humans. Most non-lethal mutants that have a non-wild type phenotype exhibit some activity, but significantly less than wild type.
In C. elegans, several mutations in ChAT have been traced to the cha-1 gene. All mutations result in a significant drop in ChAT activity. Percent activity loss can be greater than 98% in some cases. Phenotypic effects include slowed growth, decreased size, uncoordinated behavior, and lack of sensitivity toward cholinesterase inhibitors.[19] Isolated temperature-sensitive mutants in Drosophila have all been lethal. Prior to death, affected flies show a change in behavior, including uncontrolled movements and a change in electroretinogram activity.[20]
The human gene responsible for encoding ChAT is CHAT. Mutations in CHAT have been linked to congenital myasthenic syndrome, a disease which leads to general motor function deficiency and weakness. Further symptoms include fatal apnea. Out of ten isolated mutants, 1 has been shown to lack activity completely, 8 have been shown to have significantly decreased activity, and 1 has an unknown function.[21]
Clinical significance
Alzheimer's disease
The Alzheimer's disease (AD) involves difficulty in memory and cognition. The concentrations of acetylcholine and ChAT are remarkably reduced in the cerebral neocortex and hippocampus.[22] Although the cellular loss and dysfunction of the cholinergic neurones is considered a contributor to Alzheimer disease, it is generally not considered as a primary factor in the development of this disease. It is proposed that the aggregation and deposition of the Beta amyloid protein, interferes with the metabolism of neurones and further damages the cholinergic axons in the cortex and cholinergic neurones in the basal forebrain.[23]
Amyotrophic lateral sclerosis
The amyotrophic lateral sclerosis (ALS) is one of the most common motor neuron diseases. A significant loss of ChAT immunoreactivity is found in ALS.[24] It is hypothesized that the cholinergic function is involved in an uncontrolled increase of intracellular calcium concentration whose reason still remains unclear.[25]
Drugs
Neostigmine methylsulfate, an anticholinesterase agent, has been used to target ChAT. In particular, use of neostigmine methylsulfate has been shown to have positive effects against congenital myasthenic syndrome.[26]
Exposure to estradiol has been shown to increase ChAT in female rats.[27]
See also
References
- 1 2 3 GRCh38: Ensembl release 89: ENSG00000070748 - Ensembl, May 2017
- 1 2 3 GRCm38: Ensembl release 89: ENSMUSG00000021919 - Ensembl, May 2017
- ↑ "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ↑ "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ↑ Strauss WL, Kemper RR, Jayakar P, Kong CF, Hersh LB, Hilt DC, Rabin M (February 1991). "Human choline acetyltransferase gene maps to region 10q11-q22.2 by in situ hybridization". Genomics. 9 (2): 396–8. doi:10.1016/0888-7543(91)90273-H. PMID 1840566.
- 1 2 3 Nachmansohn D, Machado AL (1943). "The Formation of Acetylcholine. A New Enzyme: Choline Acetylase". J. Neurophysiol. 6 (5): 397–403. doi:10.1152/jn.1943.6.5.397.
- ↑ Berman R, Wilson IB, Nachmansohn D (September–October 1953). "Choline acetylase specificity in relation to biological function". Biochimica et Biophysica Acta. 12 (1–2): 315–24. doi:10.1016/0006-3002(53)90150-4. PMID 13115440.
- ↑ Lipmann F, Kaplan NO (1946). "A Common Factor in the Enzymatic Acetylation of Sulfanilamide and of Choline". J. Biol. Chem. 162 (3): 743–744. doi:10.1016/S0021-9258(17)41419-0.
- ↑ Lipton MA (1946). "Mechanism of the enzymatic synthesis of acetylcholine". Fed. Proc. 5 (1 Pt 2): 145. PMID 21066687.
- ↑ Nachmansohn D, Berman M (1946). "Studies on choline acetylase; on the preparation of the coenzyme and its effect on the enzyme". J. Biol. Chem. 165 (2): 551–63. doi:10.1016/S0021-9258(17)41168-9. PMID 20276121.
- ↑ Jones DH, Nelson WL (1968). "A method for isolation of coenzyme A products". Anal. Biochem. 26 (3): 350–7. doi:10.1016/0003-2697(68)90195-4. PMID 5716187.
- ↑ Govindasamy L, Pedersen B, Lian W, Kukar T, Gu Y, Jin S, Agbandje-McKenna M, Wu D, McKenna R (November 2004). "Structural insights and functional implications of choline acetyltransferase". Journal of Structural Biology. 148 (2): 226–35. doi:10.1016/j.jsb.2004.06.005. PMID 15477102.
- ↑ Oda Y (November 1999). "Choline acetyltransferase: the structure, distribution and pathologic changes in the central nervous system". Pathology International. 49 (11): 921–37. doi:10.1046/j.1440-1827.1999.00977.x. PMID 10594838. S2CID 23621617.
- ↑ Tandon A, Bachoo M, Weldon P, Polosa C, Collier B (1996). "Effects of colchicine application to preganglionic axons on choline acetyltransferase activity and acetylcholine content and release in the superior cervical ganglion". J. Neurochem. 66 (3): 1033–41. doi:10.1046/j.1471-4159.1996.66031033.x. PMID 8769864. S2CID 44586742.
- ↑ Pahud G, Salem N, van de Goor J, Medilanski J, Pellegrinelli N, Eder-Colli L (25 May 1998). "Study of subcellular localization of membrane-bound choline acetyltransferase in Drosophila central nervous system and its association with membranes". European Journal of Neuroscience. 10 (5): 1644–53. doi:10.1046/j.1460-9568.1998.00177.x. PMID 9751137. S2CID 24196247.
- ↑ Bruce G, Hersh LB (December 1987). "Studies on detergent released choline acetyltransferase from membrane fractions of rat and human brain". Neurochem Res. 12 (12): 1059–66. doi:10.1007/bf00971705. PMID 2450285. S2CID 4336737.
- ↑ Carroll PT (1994). "Membrane-bound choline-O-acetyltransferase in rat hippocampal tissue is associated with synaptic vesicles". Brain Res. 633 (1–2): 112–8. doi:10.1016/0006-8993(94)91529-6. PMID 8137149. S2CID 1139292.
- ↑ Rand, James. "Acetylcholine". WormBook.
- ↑ Rand JB, Russell RL (February 1984). "Choline acetyltransferase-deficient mutants of the nematode Caenorhabditis elegans". Genetics. 106 (2): 227–48. doi:10.1093/genetics/106.2.227. PMC 1202253. PMID 6698395.
- ↑ Greenspan RJ (1980). "Mutations of choline acetyltransferase and associated neural defects". Journal of Comparative Physiology. 137 (1): 83–92. doi:10.1007/BF00656920. S2CID 45897606.
- ↑ Ohno K, Tsujino A, Brengman JM, Harper CM, Bajzer Z, Udd B, Beyring R, Robb S, Kirkham FJ, Engel AG (13 February 2001). "Choline acetyltransferase mutations cause myasthenic syndrome associated with episodic apnea in humans". Proceedings of the National Academy of Sciences. 98 (4): 2017–2022. Bibcode:2001PNAS...98.2017O. doi:10.1073/pnas.98.4.2017. PMC 29374. PMID 11172068.
- ↑ Bartus RT, Dean RL, Beer B, Lippa AS (30 July 1982). "The cholinergic hypothesis of geriatric memory dysfunction". Science. 217 (4558): 408–14. Bibcode:1982Sci...217..408B. doi:10.1126/science.7046051. PMID 7046051.
- ↑ Geula C, Mesulam MM, Saroff DM, Wu CK (January 1998). "Relationship between plaques, tangles, and loss of cortical cholinergic fibers in Alzheimer disease". J Neuropathol Exp Neurol. 57 (1): 63–75. doi:10.1097/00005072-199801000-00008. PMID 9600198.
- ↑ Oda Y, Imai S, Nakanishi I, Ichikawa T, Deguchi T (December 1995). "Immunohistochemical study on choline acetyltransferase in the spinal cord of patients with amyotrophic lateral sclerosis". Pathol Int. 45 (12): 933–9. doi:10.1111/j.1440-1827.1995.tb03418.x. PMID 8808298. S2CID 23763400.
- ↑ Morrison BM, Morrison JH (January 1999). "Amyotrophic lateral sclerosis associated with mutations in superoxide dismutase: a putative mechanism of degeneration". Brain Res Brain Res Rev. 29 (1): 121–35. doi:10.1016/s0165-0173(98)00049-6. PMID 9974153. S2CID 28937351.
- ↑ Greer M, Schotland M (July 1960). "Myasthenia gravis in the newborn". Pediatrics. 26: 101–8. doi:10.1542/peds.26.1.101. PMID 13851666. S2CID 8672902.
- ↑ Luine VN (August 1985). "Estradiol increases choline acetyltransferase activity in specific basal forebrain nuclei and projection areas of female rats". Experimental Neurology. 89 (2): 484–90. doi:10.1016/0014-4886(85)90108-6. PMID 2990988. S2CID 1525252.
Further reading
- Oda Y (2000). "Choline acetyltransferase: the structure, distribution and pathologic changes in the central nervous system". Pathol. Int. 49 (11): 921–37. doi:10.1046/j.1440-1827.1999.00977.x. PMID 10594838. S2CID 23621617.
- Wang J, Fu X, Zhang D, Yu L, Li N, Lu Z, Gao Y, Wang M, Liu X, Zhou C, Han W, Yan B, Wang J (2017). "ChAT-positive neurons participate in subventricular zone neurogenesis after middle cerebral artery occlusion in mice". Behav. Brain Res. 316: 145–151. doi:10.1016/j.bbr.2016.09.007. PMC 5048585. PMID 27609645.
- Oda Y, Nakanishi I, Deguchi T (1993). "A complementary DNA for human choline acetyltransferase induces two forms of enzyme with different molecular weights in cultured cells". Brain Res. Mol. Brain Res. 16 (3–4): 287–94. doi:10.1016/0169-328X(92)90237-6. PMID 1337937.
- Wang J, Lu Z, Fu X, Zhang D, Yu L, Li N, Gao Y, Liu X, Yin C, Ke J, Li L, Zhai M, Wu S, Fan J, Lv L, Liu J, Chen X, Yang Q, Wang J (2017). "Alpha-7 Nicotinic Receptor Signaling Pathway Participates in the Neurogenesis Induced by ChAT-Positive Neurons in the Subventricular Zone". Transl Stroke Res. 8 (5): 484–493. doi:10.1007/s12975-017-0541-7. PMC 5704989. PMID 28551702.
- Toussaint JL, Geoffroy V, Schmitt M, Werner A, Garnier JM, Simoni P, Kempf J (1992). "Human choline acetyltransferase (CHAT): partial gene sequence and potential control regions". Genomics. 12 (2): 412–6. doi:10.1016/0888-7543(92)90395-9. PMID 1339386.
- Lorenzi MV, Trinidad AC, Zhang R, Strauss WL (1992). "Two mRNAs are transcribed from the human gene for choline acetyltransferase". DNA Cell Biol. 11 (8): 593–603. doi:10.1089/dna.1992.11.593. PMID 1388731.
- Misawa H, Ishii K, Deguchi T (1992). "Gene expression of mouse choline acetyltransferase. Alternative splicing and identification of a highly active promoter region". J. Biol. Chem. 267 (28): 20392–9. doi:10.1016/S0021-9258(19)88714-8. PMID 1400357.
- Cervini R, Rocchi M, DiDonato S, Finocchiaro G (1992). "Isolation and sub-chromosomal localization of a DNA fragment of the human choline acetyltransferase gene". Neurosci. Lett. 132 (2): 191–4. doi:10.1016/0304-3940(91)90299-9. PMID 1784419. S2CID 23030200.
- Strauss WL, Kemper RR, Jayakar P, Kong CF, Hersh LB, Hilt DC, Rabin M (1991). "Human choline acetyltransferase gene maps to region 10q11-q22.2 by in situ hybridization". Genomics. 9 (2): 396–8. doi:10.1016/0888-7543(91)90273-H. PMID 1840566.
- Viegas-Péquignot E, Berrard S, Brice A, Apiou F, Mallet J (1991). "Localization of a 900-bp-long fragment of the human choline acetyltransferase gene to 10q11.2 by nonradioactive in situ hybridization". Genomics. 9 (1): 210–2. doi:10.1016/0888-7543(91)90242-7. PMID 2004764.
- Itoh N, Slemmon JR, Hawke DH, Williamson R, Morita E, Itakura K, Roberts E, Shively JE, Crawford GD, Salvaterra PM (1986). "Cloning of Drosophila choline acetyltransferase cDNA". Proc. Natl. Acad. Sci. U.S.A. 83 (11): 4081–5. Bibcode:1986PNAS...83.4081I. doi:10.1073/pnas.83.11.4081. PMC 323670. PMID 3086876.
- Hersh LB, Takane K, Gylys K, Moomaw C, Slaughter C (1988). "Conservation of amino acid sequences between human and porcine choline acetyltransferase". J. Neurochem. 51 (6): 1843–5. doi:10.1111/j.1471-4159.1988.tb01166.x. PMID 3183663. S2CID 24613238.
- Berrard S, Brice A, Lottspeich F, Braun A, Barde YA, Mallet J (1988). "cDNA cloning and complete sequence of porcine choline acetyltransferase: in vitro translation of the corresponding RNA yields an active protein". Proc. Natl. Acad. Sci. U.S.A. 84 (24): 9280–4. doi:10.1073/pnas.84.24.9280. PMC 373252. PMID 3480542.
- Chireux MA, Le Van Thai A, Weber MJ (1995). "Human choline acetyltransferase gene: localization of alternative first exons". J. Neurosci. Res. 40 (4): 427–38. doi:10.1002/jnr.490400402. PMID 7616604. S2CID 42856768.
- Bausero P, Schmitt M, Toussaint JL, Simoni P, Geoffroy V, Queuche D, Duclaud S, Kempf J, Quirin-Stricker C (1993). "Identification and analysis of the human choline acetyltransferase gene promoter". NeuroReport. 4 (3): 287–90. doi:10.1097/00001756-199303000-00015. PMID 7682855.
- Quirin-Stricker C, Nappey V, Simoni P, Toussaint JL, Schmitt M (1994). "Trans-activation by thyroid hormone receptors of the 5' flanking region of the human ChAT gene". Brain Res. Mol. Brain Res. 23 (3): 253–65. doi:10.1016/0169-328X(94)90232-1. PMID 8057782.
- Erickson JD, Varoqui H, Schäfer MK, Modi W, Diebler MF, Weihe E, Rand J, Eiden LE, Bonner TI, Usdin TB (1994). "Functional identification of a vesicular acetylcholine transporter and its expression from a "cholinergic" gene locus". J. Biol. Chem. 269 (35): 21929–32. doi:10.1016/S0021-9258(17)31734-9. PMID 8071310.
- Kengaku M, Misawa H, Deguchi T (1993). "Multiple mRNA species of choline acetyltransferase from rat spinal cord". Brain Res. Mol. Brain Res. 18 (1–2): 71–6. doi:10.1016/0169-328X(93)90174-N. PMID 8479291.
- Misawa H, Matsuura J, Oda Y, Takahashi R, Deguchi T (1997). "Human choline acetyltransferase mRNAs with different 5'-region produce a 69-kDa major translation product". Brain Res. Mol. Brain Res. 44 (2): 323–33. doi:10.1016/S0169-328X(96)00231-8. PMID 9073174.
- Lönnerberg P, Ibáñez CF (1999). "Novel, testis-specific mRNA transcripts encoding N-terminally truncated choline acetyltransferase". Mol. Reprod. Dev. 53 (3): 274–81. doi:10.1002/(SICI)1098-2795(199907)53:3<274::AID-MRD3>3.0.CO;2-8. PMID 10369388. S2CID 39464614.
- Sakakibara A, Hattori S (2000). "Chat, a Cas/HEF1-associated adaptor protein that integrates multiple signaling pathways". J. Biol. Chem. 275 (9): 6404–10. doi:10.1074/jbc.275.9.6404. PMID 10692442.
External links
- Choline+Acetyltransferase at the U.S. National Library of Medicine Medical Subject Headings (MeSH)
PDB gallery | |
|---|---|
|